Влияние непрерывных низкочастотных и среднечастотных тональных шумов на кольчатую нерпу
С середины 20-го века уровень зашумленности мирового океана начал расти. Что было связано с техногенной деятельностью человека. Так по разным оценкам доля техногенного шума в океане составляет от 70 до 90 %. Мировой океан является пристанищем для более чем миллиона видов и общая биомасса оценивается примерно в миллиард тонн. Убедительно доказано влияние шумов и различных звуковых сигналов на китообразных. Так как у китообразных звуковая рецепция играет доминирующую роль над остальными органами чувств, то акустические помехи, генерируемые различными производственными и военными объектами, вызывают сбои в работе слухового анализатора китообразных при эхолокации и внутривидовом общении (ссылка). Так же существуют доказательства негативного влияния шумов на жизнедеятельность некоторых видов рыб и других морских обитателей (ссылка). Однако о влиянии техногенных шумов на ластоногих известно мало и данные зачастую противоречивы. Ластоногие, в отличии от других обитателей океана, не являются полностью водными животными и часть времени проводят на суше, где они спариваются, линяют и растят потомство. Что и влечет за собой отличное от других млекопитающих строение слуховой системы. Так в водной среде тюлени используют одни механизмы звуковосприятия, а на суше другие. Слуховая система ластоногих, в отличии от наземных млекопитающих, обитающих при относительно постоянном давлении, подвержена большим и резким перепадам давления при нырянии на глубину. Несомненно, что на тюленя могут оказывать негативное воздействие резонансные частоты, при которых возникает резонанс различных органов и тканей (ссылка).
1. Строение слухового аппарата ластоногих
Для всех млекопитающих характерно схожее строение слуховой системы. Но в процессе филогенеза система приобретает особенности, помогающие виду приспособиться к среде обитания. У ластоногих, в результате перехода к полуводному образу жизни слуховая система приобрела ряд анатомических изменений.
Самые кардинальные перестройки у морских млекопитающих приобрели периферические отделы и начальные звенья центрального слухового пути. Именно эти отделы наиболее подвержены филогенетическим изменениям в процессе адаптации к факторам окружающей среды.
У всех млекопитающих периферический отдел слуховой системы включает в себя наружное, среднее и внутреннее ухо.
У морских млекопитающих именно наружное ухо претерпело наибольшие изменения (Солнцева 1974; Богословская, Солнцева 1979). У наземных млекопитающих Ушная раковина хорошо развита и у многих видов имеет довольно сложное строение, слуховой канал короткий и прямой. Некоторые полуводные животные имеют специальные кожные выросты на ушной раковине, которые при погружении в воду закрывают ушной канал. У таких животных как сивучи и морские котики, которые проводят в воде больше времени, ушные мышцы на воздухе раздвигают складки ушной раковины, а в воде снова сдвигают, препятствуя попаданию жидкости в слуховой канал. У настоящих тюленей, проводящих в воде более времени, чем на суше, ушная раковина редуцирована и функцию защиты играет слуховой канал. Он сильно удлинен, имеет S-образную форму и, в отличии от наземных млекопитающих, располагается непосредственно под кожей. Слуховой канал у настоящих тюленей способен к изменению диаметра под давлением воды или при сокращении мышц (King, 1964; Dehnhardt, 2002).
В месте коленного изгиба проход сужается, в проксимальной части расширяется и приобретает форму овала.
Так же у ластоногих присутствует еще целый ряд адаптационных изменений наружного уха. Подковообразный наружный хрящ, окружающий наружный слуховой проход у наземных млекопитающих, видоизменен в четыре подвижно соединенными друг с другом пластины, способные изменять диаметр и форму просвета слухового канала (Солнцева, 1973; Rampsrashad ets. al., 1973). В костном отделе слухового канала располагаются хорошо развитые венозные сосуды.
Среднее ухо также имеет анатомические особенности. У морских млекопитающих барабанная полость схожа с барабанной полостью других млекопитающих, способных к ультрозвуковой ориентации и эхолокации (летучие мыши, землеройки) и имеет сферическую форму (Богословская, Солнцева, 1979). У настоящих тюленей барабанная полость выстлана толстой слизистой оболочкой. Также хорошо развиты ушные мышцы, они имеют вид коротких, но широких пучков, что тоже характерно для животных использующих эхолокацию. слуховые косточки соеденены жестко, в отличии от наземных млекопитающих. Весовые отношения молоточка и наковальни, относительные размеры слуховых косточек и их оси вращения также сильно отличаются от отношений наземных млекопитающих. Что повлекло за собой изменения коэффициента передачи звукового давления. Так у наземных и полуводных форм коэффициент составляет 25-29, у ушастых и настоящих тюленей 40-60 (Солнцева, 1973). Подобные анатомические особенности повышают коэффициент передачи звукового давления в среднем ухе, что позволяет тюленям более эффективно ориентироваться в воде, а также расширяет диапазон воспринимаемых частот.
В отличии от наземных млекопитающих, у которых улитка имеет до 5 оборотов, у тюленей улитка имеет только 2-2.5 оборота, причем наиболее развит базальный оборот. Стенка между базальным и следующим оборотом улитки сильно утолщена. Расстояние между спиральными пластинками узкое.
Отличия от наземных млекопитающих наблюдаются также в слуховых центрах мозга, особенно в начальных звеньях центрального акустического пути. Слуховой нерв представлен в виде сложной волокнистой системы и , если у наземных млекопитающих имеет толщину 2-4 мкм, то у морских млекопитающих толщина достигает 5-7 мкм, причем имеет неоднородную толщину. Что позволяет менять частоту проводимых импульсов и активно влиять на характер передаваемой информации.
Акустическое общение является наиболее эффективной формой коммуникации при обитании в укрытиях, в водной среде, для осуществления связи в ночное время и в сильно загрязненных водоемах, где коммуникация на посредством фото или хеморецепции затруднено или невозможно.
Для ластоногих слух играет важнейшую роль для пространственной ориентации и коммуникации, и по значимости превосходит зрение и хеморецепцию (Вуд, 1979; King, 1964). Так зафиксированы случаи, когда зверобои отлавливали хорошо упитанных полностью слепых тюленей. Также наблюдения в неволе показали, что поведение слепых ластоногих мало чем отличается от зрячих. Что свидетельствует о том, что зрение не играет основной роли при охоте и не является основной рецептивной системой (King, 1964; Dehnhardt, 2002).
Действительно, на глубинах, где в основном ластоногие охотятся освещенность крайне низкая, либо отсутствует полностью, так что использование зрения становиться невозможным. В тоже время звук в воде распространяется в 5 раз быстрее, чем на воздухе, и распространяется на большие расстояния. Поэтому использование слуха для ориентации в водной среде более выгодно. Однако ориентировка по свехнизким и низким частотам, из-за большой длины волны, затруднено, поэтому у ластоногих частотный диапазон расширен за счет высоких частот и повышена оптимальная частота слуха. Достоверных сведений о способности ластоногих воспринимать инфразвуки нет.
Также нет достоверных данных о использовании ластоногими активной эхолокации. До середины 1960-х годов никто не занимался эхолокацией у ластоногих, так как не было замечено, чтобы ластоногие производили какие либо звуки под водой. Но в 1965 году американский акустик Т. Полтер установил, что во время поиска рыбы морские львы испускали группы коротких импульсов – до 30 секунд, длительность каждого импульса составляла 3-5 мс, с частотами от 3 до 13 кГц. Слепой морской лев следовал вдоль неровной стены, при этом лая, причем темп лая изменялся если что либо встречалось на пути. Предположение, что настоящие тюлени также используют этот механизм, было выдвинуто зоологами доктором Дианой Ненуф и доктором М. Бенджамином Девисом из Мемориального университета Ньюфаундленда (Канада). Записи голоса обыкновенного тюленя (Phoca vitulina) в неволе и на воле показали, что эти виды используют эхолокацию, когда отсутствуют зрительные стимулы, продуцируя, как минимум, два типа сигналов (Шукер, 2006). Кольчатая нерпа, животное с точки зрения эхолокации весьма перспективное, оказалась способной щебетать, рычать, лаять и тявкать под водой. Однако, скорее всего, все эти звуки используются исключительно для общения (Сергеев, 1980). Таким образом, до настоящего времени вопрос о том, способны ли ластоногие к эхолокации, в достаточной степени не выяснен. Так, подводные крики гренландского тюленя и тюленя Уэдделла некоторые исследователи рассматривают как локационные посылки (Terhune, 2001), а исследования Р. Шустермана и соавторов (Schusterman et al., 2000) свидетельствуют об обратном. Непосредственное изучение реакции мозга морских львов на звук показало, что их слуховые центры не так хорошо, как у дельфинов, приспособлены для анализа коротких звуков, которые обычно используются для локации. По всей видимости, ластоногие способны к эхолокации, но в естественных условиях редко ей пользуются, полагаясь при поиске пищи даже в мутной воде лишь на пассивную локацию, ориентируясь на звуки, издаваемой потенциальной добычей. С точки зрения использования эхолокации при питании, можно считать наиболее продвинутыми арктических тюленей, которым приходится добывать пищу на акватории, покрытой льдом в условиях полярной ночи. Так некоторые специалисты считают, что гренландские тюлени в своем развитии прогрессировали больше, чем другие виды настоящих тюленей, и используют эхолокацию во время миграций на большие расстояния и охоты на больших глубинах. Опыты Л.А. Попова показали, что гренландские тюлени хорошо разбираются в голосах моря и могут ловить добычу на слух в полной темноте, лоцируя объект и находя его по отраженным сигналам (Стюарт, 1978).
2. Характеристики шумов
Звуковой сигнал можно представить, как совокупность различных синусоидальных составляющих. Каждая составляющая характеризуется рядом параметров.
Высота звука - определяется частотой звуковой волны (или, периодом волны). Чем выше частота, тем выше звучание.
Высота звука измеряется в герцах (Гц, Hz) или килогерцах (КГц, KHz). 1 Гц = 1/С. То есть колебание в 1 Гц соответствует волне с периодом в 1 секунду.
Громкость звука - определяется амплитудой сигнала. Чем выше амплитуда звуковой волны, тем громче сигнал.
Громкость звука измеряется децибеллах и обозначается дБ. Белл определяется, как логарифм отношения электрических, акустических или других мощностей по формуле Бел = log(P1/P0).
Согласно закона Вебера-Фехнера, чувствительность уха к громкости звука носит логарифмической характер, поэтому их мощность, выраженная в децибелах, точнее отражает восприятие звуков человеком и животными.
Громкость — это уровень мощности, которая пропорциональна амплитуде звукового сигнала. Громкость определяют в дБm — относительно стандартного значения 1 мкПа. Тогда шкала приобретает абсолютное значение. Также, часто громкость определяют относительно порога слышимости человеческого уха 22 мкПа.
Субъективно ухо воспринимает не мощность, а звуковое давление на барабанную перепонку. Мощность звука — это совокупная звуковая энергия, которую излучает источник звука, например громкоговоритель. Звуковое давление — это звуковая энергия, которая попадает на единицу площади, удаленную от источника звука на расстояние 1м.
Двухкратное увеличение звукового давления соответствует 6 дБ, а десятикратное — 20 дБ.
Звуки, которые различаются на 3 дБ, ухом воспринимаются одинаковыми по громкости. Звук, давление которого на 10 дБ выше, для уха будут звучать в два раза громче. Следует отметить, что громкость — субъективная характеристика, которая зависит от частоты. Лучше всего ухо человека воспринимает звук в диапазоне от 1 до 4 кГц. За 0 дБ принят уровень звукового давления, соответствующий порогу слышимости здорового молодого человека в этом диапазоне частот.
Стоит отметить, что ухо человека воспринимает одинаковую громкость на разных частотах, как звуки разной громкости.
Понятие "шум" весьма субъективно. Всякий нежелательный в данный момент звук (или звуки) человек воспринимает как шум. Одни и те же звуки разными людьми могут восприниматься по-разному. Физиологи и гигиенисты определяют шум как звук, оцениваемый негативно и наносящий вред здоровью. Машины и механизмы, используемые на производстве, являются источниками звуков различной частоты и интенсивности, изменяющихся во времени. Поэтому производственный шум рассматривают как совокупность звуков различной интенсивности и частоты, беспорядочно изменяющихся во времени и вызывающих у работающих неприятные субъективные ощущения.
По частотным характеристикам акустические шумы подразделяются на инфразвуковые или сверхнизкочастотные с частотами ниже 20 Гц, низкочастотные (20-300 Гц), среднечастотные (300-800 Гц), высокочастотные (800-20000 Гц) и ультразвуковые или сверхвысокочастотные (20-150 кГц). По спектральным характеристикам разделяют широкополосные с непрерывным спектром шире 1 октавы и тональные шумы с ярко выраженными дискретными тонами. По временным характеристикам шумы делятся на постоянные, когда уровень шума меняется не более чем на 5 дБ, и непостоянные. Которые, в свою очередь подразделяются на колеблющиеся, когда уровень постоянно изменяется во времени; прерывистые, когда уровень шума меняется ступенчато не более чем на 5 дБ, с длительностью интервала более секунды; импульсные, состоящих из нескольких звуковых сигналов с интервалами менее секунды.
3. Воздействие различных шумов на ластоногих
Ластоногие способны воспринимать звуки в широком диапазоне. Нижним пределом является частоты 16-20 Гц, которые тюлени одинаково хорошо воспринимают как в воде, так и на суше. На суше параметры слуха у тюленей схожи с человеческими и звуки частотой выше 20 кГц тюлени воспринимают плохо. Но при погружении частотный диапазон расширяется до 55-60 кГц. Также смещается диапазон частот наилучшей восприимчивости, так у кольчатой нерпы он составляет 1-45 кГц, у обыкновенного тюленя 1-32 кГц, у гренландского тюленя 2-23 кГц (King, 1964). Именно на данных частотах, из-за короткой длины волны, возможно точное ориентирование на источник звука, которым может быть объект охоты, сородич, либо хищник или иная угроза. Поэтому для тюленей представляет опасность шумы именно на этих частотах. Однако доля звуков данных частот в техногенных шумах невелика, а также звуки на высоких и ультразвуковых частотах обладают значительно меньшей энергией, чем инфразвуки и звуки низких и средних частот, и поэтому распространяются на значительно меньшие расстояния.
Главными источниками высокочастотных и ультразвуковых сигналов являются сонары и гидролокаторы, их излучение характеризуется высокой мощностью, и вблизи источника представляет угрозу для любого организма. Особенно опасны звуки высокочастотных и ультразвуковых диапазонов для китообразных, которые используют эхолокацию. Данные акустические сигналы, во-первых, глушат животных, а во-вторых, создают помехи в работе эхолокационной системы, делая невозможным различать окружающие предметы. Однако, в отличие от китообразных, ластоногие обладают не столь чувствительным слухом, а также не пользуются активной эхолокацией (по крайней мере, эхолокация не доминирует над другими органами чувств у тюленей). Так существует мнение, что способность тюленей различать ультразвуки, является оборонительной адаптацией, против естественных врагов – касаток, которые активно используют эхолокацию (King, 1964).
Часть спектра высоких частот (от 800 до 9000 Гц) присутствует в шуме, создаваемом судами (без учета работы эхолотов и сонаров) и промышленными объектами. Реже в данном шуме присутствуют звуки с частотами выше 10 кГц. Данные звуки характеризуются низкой интенсивностью и распространяются на небольшие дистанции. Однако среднечастотные, низкочастотные и инфразвуковые составляющие подобных шумов высоки, особенно инфразвуковые и низкочастотные.
Уровни шума внутри судна и наведённые вибрации корпуса делают необходимым рассматривать его как достаточно мощный источник высокоинтенсивного акустического излучения в окружающую водную среду (ссылка).
Коммерческий флот является главным источником низкочастотных звуков (5 - 500 Гц) в мировом океане. Проходящие в отдалении суда увеличивают уровень фонового шума на больших площадях мирового океана. Звуки, издаваемые отдельными судами, часто невозможно различить ни во времени, ни в пространстве среди шумов, издаваемых в отдалении судами. Суда создают звуки при работе гребных винтов, энергетических установок и благодаря гидравлическим потокам воды, омывающей корпус корабля. В целом суда издают целый ряд шумов в частотах от 10 Гц до 10 кГц. Последние исследования уровней шумов, издаваемых малыми скоростными катерами, показывают, что уровни пиков их спектральной плотности находятся в диапазоне 350-1 200 Гц и 145-150 дБ на расстоянии 1 м (Bartlett and Wilson, 2002). Richardson et. al. (1995) указывают в своей работ результаты замера уровня шумов в 162 дБ на частоте 630 Гц (на расстоянии 1 м), производимых буксиром и баржей, двигающимся со скоростью 18 км/час, а также уровни шума, производимого большим танкером: около 177 дБ (на расстоянии 1 м) 1/3 октавной полосе с центральной частотой 100 Гц. Большие суда имеют более мощные двигательные установки и их моторы и гребные винты медленнее вращаются. Большая площадь корпуса судна более эффективно проводит шум от работы механизмов через обшивку в морскую среду. Поэтому практика показывает, что чем больше судно, тем выше уровень производимых шумов и тем ниже преобладающий диапазон частот звуков. К тому же при увеличении скорости движения уровень интенсивности производимых шумов на любом конкретном судне увеличивается.
Уровень шума среднеоборотных дизелей на частотах порядка 500 Гц достигает величины 100 дБ. По данным лаборатории «Noise» CIMAC, сотрудниками которой были проведены исследования 200 типов дизелей мощностью от 8 до 6000 кВт с частотой вращения коленчатого вала от 5 об/с до 50 об/с, из общего числа обследованных устройств, 70% имели максимальный уровень шума 95 − 105 дБ, а у отдельных образцов величина превышала болевой порог и составляла 110 дБ. Основными источниками шума и вибраций на судах всех типов являются их силовые установки, механические передачи и гребные винты. Каждый из этих источников помимо воздушных и гидродинамических шумов способствует возникновению вибраций судовых конструкций, которые в свою очередь превращаются во вторичные излучатели. Судовые шумы и вибрации возникают при действии на конструкции динамических сил. Среди многообразия типов динамических сил можно выделить несколько основных.
Первую группу составляют силы механического происхождения, возникновение которых вызвано неуравновешенностью вращающихся частей механизмов, асимметричностью жёсткости роторов, технологическими и сборочными дефектами механической обработки соприкасающихся деталей, а также возмущениями вследствие соударения деталей в поршневых и кривошипно-шатунных механизмах.
Во вторую группу целесообразно объединить силы электромагнитного происхождения, вызываемые нестационарными магнитными полями в электрических машинах. Переменные во времени магнитные поля вызывают колебания элементов электрических машин, в частности, вследствие магнитострикционного эффекта.
Третья группа сил обусловлена аэро- и гидродинамическими явлениями, наблюдающимися в насосах, турбинах, компрессорах, трубопроводах, запорной арматуре и т. п.
Все три типа сил, в конечном счете, по отношению к элементам конструкций, способных совершать колебания, являются внешними возмущающими. Колебания конструкций становятся возможными при наличии у них упругих свойств, которые формируют возвращающие силы. В общем случае элемент судовой конструкции обладает, как и всякое свободное твёрдое тело шестью степенями свободы. К таким системам относятся устройства, соединённые с палубой или переборками через виброизолирующие конструкции.
Наиболее интенсивные акустические шумы обусловлены работой судового движителя и сопровождающими работу винтов гидродинамическими явлениями. Гребной винт вследствие конечного числа лопастей создаёт в жидкости периодические разряжения и сжатия, частота которых определяется в виде произведения угловой скорости вращения на число лопастей. Изменение давления в жидкости вследствие её сжимаемости приводит к возникновению волн акустического диапазона, именуемых звуком вращения гребного винта. Спектр шума имеет явно выраженные максимумы в низкочастотной области, вызванные периодическим вытеснением жидкости из области вращения винта и высокочастотную составляющую, турбулентного происхождения.
Максимальный уровень шума в воде наблюдается в области частот 250 − 450 Гц, причём выше 0, 8 кГц уровень падает со скоростью 10 дБ/окт.
В случае движущегося судна можно выделить три характерных вида высокоинтенсивных шумов: работающие судовые механизмы, основные и вспомогательные; гребной винт; гидродинамические шумы турбулентного происхождения; кавитационный шум, обусловленный разрывами сплошности воды, как правило, на кромках лопастей гребного винта; шумы, генерируемые носовым и кормовым бурунами.
Также источником подводных и надводных шумов являются буровые установки и судна обеспечения приписанные к ним. Одним из интенсивных источников шума на морских нефтегазопромысловых сооружениях являются приводные двигатели различного назначения. Основными источниками шума на подвышечном портале плавучей буровой установки и бурового судна являются: буровая лебедка, ротор, двигатели привода лебедки и ротора, автоматический буровой ключ, исполнительные механизмы системы управления буровыми механизмами, пневмокраны и др. На рабочей площадке буровых морских стационарных платформ кроме перечисленных источников размещены механизмы передачи мощности от расположенных на площадке силовых дизельных агрегатов или электродвигателей, также создающие шум и вибрацию. Уровни шума в рабочей зоне буровых платформ достигают 107 дБ. В связи с наличием большого числа источников шума в различных точках площадки уровни шума разнятся на 4–7 дБ. Спектр шума зависит от количества и состава включаемых в работу источников, кроме того, шумовая картина определяется видом технологического процесса.
Так, например, акустические измерения показали, что платформа «Моликпак» и нефтедобывающий комплекс «Витязь» излучают тональные и узкополосные акустические сигналы, в том числе на частотах меньше 30 Гц, спектральный уровень, которых на 10-15 дБ выше, чем уровень широкополосного шума. Было установлено, что наиболее интенсивными источниками акустического шума с частотой менее 1 кГц являются суда обеспечения. Спектральные уровни этих тональных и узкополосных сигналов (с частотой менее 500 Гц) составляли 80-90 дБ (ссылка).
Но, несмотря, на генерируемые судами и промышленными объектами интенсивные шумы, ластоногих часто замечают вблизи буровых станций, портов и доков, где они охотятся, а также, иногда отдыхают вблизи. В некоторых странах даже существуют «тюленьи бомбы», которыми с рыболовецких судов отпугивают тюленей, охотящихся на промысловую рыбу, непосредственно вблизи судна. Если при первых взрывах, животные уходили и возвращались только через несколько минут, то после 3-4 взрыва, перестали них реагировать. Также тюлени спокойно существуют вблизи аэродромов и шум взлетающего реактивного самолета (130дБ на расстоянии 100м, после прохождения границы воздух-вода 100дБ на глубине 1м) (ссылка).
На данный момент не зарегистрировано ни одного случая гибели тюленя от воздействия промышленного шума, животные гибнут от химического загрязнения, а также от механического воздействия (попадают в работающие винты, запутываются в сетях и тралах, бьются о корпус судов). Однако очень мало известно о последствиях долговременного воздействия промышленных шумов на организм тюленя. О характере данных воздействий можно судить по влиянию шумов на человека и других наземных млекопитающих.
Установлено, что восьмичасовое воздействие шума интенсивностью 90 дБ приводит к тугоухости 5% подвергающихся шумовому воздействия. При уровнях шума 95 Дб процент частичных поражений слуха составляет уже 10%. Негативное воздействие акустического шума наблюдается не только на органы слуха. Дело в том, что волокна слуховых нервов достигают центральных областей продолговатых областей мозга, откуда раздражения в виде электрических импульсов распространяются далее. Эти важные области мозга соседствуют с центрами управления мускулами век, дыханием, процессом кровообращения, координируют реакции. Другими словами, изолированные раздражения одного органа влекут за собой комплексную реакцию организма. Влияние шума на вегетативную нервную систему наиболее выпукло проявляется на изменении функционирования системы кровообращения, увеличение артериального давления и частоты сердечных сокращений, обусловленных сокращением ударного объёма сердца и повышением периферического сопротивления кровеносных сосудов. При воздействии высокоинтенсивного шума наблюдается расширение зрачков, что вызывает уменьшение глубины резкости (ссылка). По-видимому, для ластоногих реальную опасность представляют либо длительное воздействие шумов, в результате которых снижается чувствительность органа слуха, либо воздействие резонансных частот высокой мощности, которые вызывают резонанс внутренних органов.
Терпимость ластоногих к высокому уровню подводных шумов изучена недостаточно и точных данных нет. Однако, на основе данных целого ряда исследований предсказывается, что воздействие шумов с уровнем приблизительно 140 дБ относительно 1 мкПа может вызвать у ластоногих временную потерю слуха и вряд ли морские млекопитающие (включая и тюленей) будут оставаться в зоне облучения шумами с уровнем 120-140 дБ относительно 1 мкПа достаточно долго, чтобы пострадать от временной или, возможно, постоянной потери слуха. Полезная аналогия по возможной ситуации в проливе Лаперуза представлена в работе по воздействию подводных шумов от круизных судов на сивучей и других морских млекопитающих, обитающих в национальном парке Глэсиер-Бей на Аляске. Эта работа была поведена для оценки возможных изменений в движение этих судов (NPS 2003). На основе шумовых характеристик судов было рассчитано, что круизные суда, идущие со скоростью 10 узлов излучают шум с уровнем 130 децибел и выше (интенсивность в 130 дБ была выбран как уровень, при котором морские млекопитающие могут реагировать на звук) на расстояние около 500 метров (LGL 2003, согласно ссылке в NPS 2003). На основе образца звуковой сигнатуры круизного судна, идущего со скоростью 19 узлов (195 дБ относительно 1 мкПа на расстоянии 1 м), в работе LGL предсказано, что в заливе Глэсиер-Бей оно будет излучать шум на уровне 130 дБ и выше на расстояние до 5 км (в радиусе от судна). Для сравнения, большие танкеры обычно производят подводные шумы с интенсивностью 175-195 дБ относительно 1 мкПа на расстоянии 1 м ((Richarsdson et al. 1995; Hildebrand 2004). Хотя прямое сравнение и невозможно из-за различных географических особенностей, можно предположить, что для танкеров, идущих по проливу Лаперуза, будут иметь место аналогичные зоны шумового облучения. Следует также учитывать, что движущееся судно не является постоянным источником шума для неподвижного объекта. Согласно оценкам с использованием данных исследований в заливе Глэсиер-Бей, максимальное время воздействия шумов интенсивностью 130 дБ или выше от судна, движущегося со скоростью 19 узлов, на неподвижный объект будет около 17 минут (NPS 2003). Это время меньше продолжительности воздействия шумов в 20-22 минуты, которая вызывает временное снижение слуховой чувствительности (временное смещение порога) у пятнистых тюленей, морских слонов и калифорнийских тюленей (Kastak et al, 1999).
Для выявления влияния шумов на кольчатую нерпу были выбраны 2 типа постоянных детерминированных тональных шумов, которые оказывают наиболее раздражающее воздействие на человека и некоторых наземных млекопитающих (Глебова, 2007). Первый тип по звучанию напоминал сирену: частота звучания линейно возрастала с нижнего значения диапазона до верхнего за 1 минуту, затем цикл повторялся. Второй тип представляет из себя звучание на случайной частоте, входящей в заданный диапазон. Для оценки величины влияния шума на организм кольчатой нерпы использовалось изучение изменений в двигательной активности и пищевой мотивации животного.
Чтобы как можно больше изолировать экспериментальное животное от воздействия посторонних шумов, неизбежно возникающих на акватории Кольского залива, где находиться вольер животного, решено было поместить нерпу в бассейн, находящийся в помещении, где уровень шумового фона намного ниже, чем в вольере.
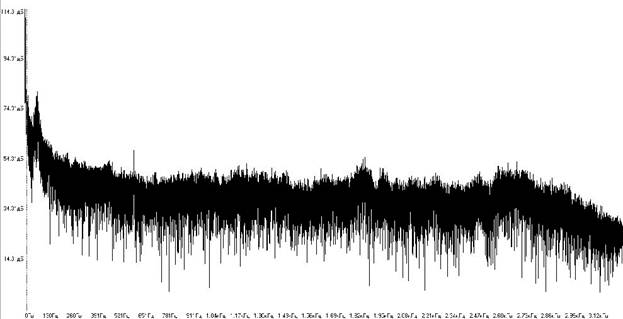
Рисунок 1. Спектр шума на акватории Кольского залива вблизи губы Тюва.
Из рисунка 1 видно, что на акватории уровень подводного шума составляет 56-70 дБ, причем максимум приходиться на сверхнизкие частоты (до 100-140 дБ), уровень низких составляет 70-80 дБ и после 300-350 Гц постепенно снижается до значений в 25-30 дБ.
В помещении, где проходил эксперимент, находились: бассейн размерами 2х2х1м., установка для генерации звукового сигнала, видеофиксации поведения животного и записи шумового фона.
Установка состояла из: ноутбука, на котором были установлены программы по генерации звука (SweepGen 3.5.2), программа для видеонаблюдения (WebCanMonitor 5.2), программа для записи и анализа акустического сигнала (Анализатор Сигнала 2007); веб-камеры Genius e-messenger 112; гидроакустической антенны ЦГП-4а и цифрового модуля БВС-ЦМ; акустического усилителя и широкополосных колонок.
При помощи программы SweepGen генерировались 2 типа непрерывных детерминированных тональных шумов: линейное возрастание частоты звучания и звучание случайной частоты в заданном диапазоне. Усилитель был подключен к аудио-выходу ноутбука и усилив сигнал, передавал его на колонки. В тоже время круглосуточно велось видеонаблюдение за животным с помощью веб-камеры и программы WebCanMonitor, которая позволяла создавать видеофайлы продолжительностью по 10 минут. Для удобства анализа, а также экономного расходования дискового пространства, видео записывалось в avi формате с кодеком видеосжатия DivX® 6.8.2 Codec (2 Logical CPUs), при разрешении 320 на 240 точек, с цветопередачей в градациях серого. Четыре раза в сутки производилась минутная запись шумового фона с помощью гидроакустической антенны ЦГП-4а. Что позволяло выявить частотные и мощьностные характеристики шумового фона (в соответствии с СИ использовались децибелы, рассчитанные относительно давления в 1 мкПа).
Для того чтобы во время эксперимента вода не попала на работающую электротехнику, приборы были покрыты чехлами из полипропилена, а компьютер был огорожен занавесью, также из полипропилена.
Для тестов на пищевую мотивацию и общее состояние животного использовались объекты, изображенные на рисунке 2. Двигательная активность изучалась с помощью видеозаписи, а пищевая мотивация с помощью тестовых команд, предъявляемых во время кормления. Кормление проходило 4 раза в сутки в 9:00, 15:00, 21:00 и 3:00, за время кормления скармливалось 200г. мойвы, единовременное подкрепление составляло 25-30г., то есть 1 рыбка. Тестовые команды включали в себя команды на стабильность и моторную функцию. Первая команда была «таргет»: по команде животное должно было удерживать таргет вплоть до подачи бридж-сигнала (свистка), в случае верного выполнения животное получало поощрение (кусочек рыбы весом 25-30г.). Вторая команда «ищи»: животное также удерживает таргет, затем подается команда «ищи» (жестом или голосом), животное должно найти и обозначить касанием объект (бутылочка из-под йогурта, подвешенная у края вольера). Данные команды чередовались в псевдослучайной последовательности (не более 2 одинаковых команд подряд), для того чтобы животное не могло распознать зависимость и выполняло именно подаваемые команды. Так же критерием служил факт того берет или не берет нерпа корм из рук экспериментатора.
Для изучения двигательной активности, в ходе анализа отснятого видеоматериала, были выделены наиболее характерные спокойные элементы двигательной активности животного (см. рисунок 3):
Нерпа «висит» в центре бассейна, голову держит под водой и периодически поднимает ее для вдоха (а)
Медленно плавает кругами в направлении по часовой стрелке (б)
Неспешно «дрейфует» по бассейну, изредка подгребая передними ластами (в)
Пытается заглянуть за бассейн у стенки (г)
Скребется когтями передних ласт о дно бассейна (д)
Тянется к сетке (е)
Также выделены элементы, характеризующиеся как возбужденные:
Быстро плавает кругами в направлении по часовой стрелке, при этом движения резкие (ж)
Быстро плавает кругами в направлении против часовой стрелки, при этом движения резкие (з)
Животное подходит к углу, а затем по диагонали, рывком перемещается в другой угол, без переворота вдоль горизонтальной оси (и)
Животное подходит к углу, а затем по диагонали, рывком перемещается в другой угол, с переворотом вдоль горизонтальной оси (к)
Частоту дыхания возможно было оценить лишь тогда, когда животное либо «висело» в центре бассейна, либо когда «дрейфовало». Так как в основном нерпа предпочитала держать голову под водой, то количество подъемов головы из воды и считалось за частоту дыхания. Так как животное порой «висело» по 5-8 минут, представлялось возможным довольно точно определить данный параметр.
Стоит отметить, что при содержании в вольере нерпа четко, быстро и без сбоев выполняла команды «таргет» и «ищи», причем, данные команды подавались как жестом, так и голосом. Также нерпа безбоязненно брала рыбу из рук тренера и разрешала себя погладить, то есть допускала тактильный контакт, что говорит о том, что животное не видела в тренере опасности.
До того как нерпу поместили в бассейн, были сняты показания уровня шума в бассейне. Общий уровень составил 35-40 дБ. На низких частотах достигал уровня 56-60 дБ (см. рисунок 4).
После помещения животного в бассейн общий уровень шума вырос на 5-10 дБ и составил 40-50 дБ, по частотным характеристикам изменился не сильно. Животное было помещено в бассейн в 13:25. В течение суток животное адаптировалось к новым условиям. Как только животное освободили из сетки, в которой его переносили, животное отплыло на середину бассейна и оттуда начало шипеть на людей, после начало бросаться на уходящих людей, при этом фыркало и било передними ластами по воде. Просматривая видеозапись периода с помещения животного в бассейн до первого кормления можно было видеть, что первые полчаса животное нервничало, постоянно скреблось о дно бассейна, пыталось вылезти, либо быстро плавало по кругу (по часовой стрелке). При первом кормлении, в 15:00 , выполнило 6 из 10 предъявлений на «таргет» и 3 из 10 на поиск предмета. Из рук экспериментатора корм брать отказывалось, держалось на расстоянии 1-1.5 метра. Животное проявляло агрессию по отношению к людям (делало рывок в сторону человека, при этом хлопая передними ластами по воде), что, вероятно, было проявлением естественной активно-оборонительной реакции свойственной кольчатым нерпам. Просмотр периода между кормлениями показал, что животное успокоилось. Нерпа либо висела в центре бассейна, изредка скреблась когтями передних ласт о дно, либо неспешно совершала 2-3 круга по часовой стрелке. Частота дыхания составляла в среднем 0.30±0.08 вдоха в минуту. На втором кормлении, 6 часов спустя (21:00), животное два раза кинулось на вошедшего человека, но больше за время кормления агрессии не проявляло. Пищевая мотивация была высока, животное выполнило все предъявления (10/10 «таргет» и 10/10 ищи), причем команды выполняло быстро и четко, не совершая лишних движений. Корм из рук в основном н