Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Міністерство освіти і науки України
Дніпродзержинський державний технічний університет
Кафедра екології та біотехнології
Дисципліна Загальна мікробіологія
КУРСОВА РОБОТА
на тему
Загальна характеристика і особливості життєдіяльності бактерій родини Enterobacteriaceae
Дніпродзержинськ
2009
ЗМІСТ
Вступ
1. Морфологія бактерій родини Enterobacteriaceae
2. Фізіологія бактерій родини Enterobacteriaceae
3. Метаболізм бактерій родини Enterobacteriaceae
4. Генетика бактерій родини Enterobacteriaceae
5. Екологія бактерій родини Enterobacteriaceae
6. Антигени бактерій родини Enterobacteriaceae
7. Патогенність і токсиноутворення бактерій родини Enterobacteriaceae
8. Резистентність бактерій родини Enterobacteriaceae
9. Патогенез бактерій родини Enterobacteriaceae
10. Імунітет до бактерій родини Enterobacteriaceae
11. Лабораторна діагностика бактерій родини Enterobacteriaceae
12. Профілактика і лікування захворювань викликаних бактеріями родини Enterobacteriaceae
13. Динаміка зміни резистентності умовно-патогенних ентеробактерій шлунково-кишкового тракту до антимікробних препаратів
Перелік посилань
ВСТУП
Важливу роль в життєдіяльності людини відіграє мікрофлора товстого кишечника – своєрідний екстракопоральний орган. Вона є антагоністом гнилісної мікрофлори, так як продукує молочну, оцтову кислоти, антибіотики і інше. Відома її роль в водно-сольовому обміні, регуляції газового складу кишечнику, обміні білків, вуглеводів, жирних кислот, холестерину і нуклеїнових кислот, а також продуктів біологічно активних сполук – антибіотиків, вітамінів, токсинів та ін. Морфокінетична роль мікрофлори складається в її участі в розвитку органів і систем організму; вона приймає участь також в фізіологічному запаленні слизистої оболонки і зміні епітелію, перетравленні і детоксикації екзогенних субстратів і метаболітів, що порівнюється з функцією печінки. Нормальна мікрофлора виконує, крім того, антимутагену роль, руйнуючи канцерогенні речовини в кишечнику. В той же час деякі бактерії можуть продукувати потужні мутагени. Пристінкова мікрофлора кишечника колонізує слизову оболонку у вигляді мікроколоній, утворюючи своєрідну біологічну плівку, яка складається з мікробних тіл і екзополісахаридного матриксу. Екзополісахариди мікроорганізмів, які називаються глікокаліксом, захищають мікробні клітини від різних фізико-хімічних і біологічних впливів. Слизова оболонка кишечника також знаходиться під захистом біологічної плівки.
Великий вплив чинить мікрофлора кишечника на формування і підтримку імунітету. В кишечнику міститься близько 1,5 кг мікроорганізмів, антигени яких стимулюють імунну систему. Природнім неспецифічним стимулятором імуногенезу є мурамілдипептид, який утворюється із мікрофлори під дією лізоциму і інших ліричних ферментів, які знаходяться у кишечнику.
Важливою функцією нормальної мікрофлори кишечнику є її участь в колонізаційній резистентності, під якою розуміють сукупність захисних факторів організму, і конкурентних, антагоністичних і інших особливостей анаеробів кишечнику, які надають стабільність мікрофлорі і запобігають колонізації слизових оболонок сторонніми мікроорганізмами (1).
До групи кишкових бактерій відносяться дуже багато бактерії, зокрема родина Enterobacteriaceae. Це велика родина яка включає такі роди як Salmonella, Escherichia, Klebsiella, Proteus, Shigella. Більшість бактерій цієї родини складають значну частину нормальної аеробної флори кишечника і зазвичай не викликають захворювань, а навпаки, беруть участь у забезпеченні його нормального функціонування. Ці мікроорганізми стають патогенними лише у випадках проникнення їх у тканини організму, особливо в сечовидільні шляхи, легені, очеревину чи мозкові оболонки, де вони здатні викликати запальний процес. Вони можуть проникати в кров'яне русло (при використанні венозних чи уретральних катетерів); попадати в сечостатеві шляхи при проведенні інструментальних досліджень (нозокоміаль-на передача); викликати інфекційні ураження сечовидільних шляхів після введення контамінованих лікарських препаратів, особливо після внутрішньовенного їхнього застосування. Можливий статевий шлях передачі.
Бактерії родини Enterobacteriaceae часто обумовлюють вторинні інфекції, у тому числі пневмонії, ендокардити (запалення внутрішньої оболонки серця), бактеріємії, особливо в людей, що приймають наркотики, і в пацієнтів клінік.
Прояви інфекцій, викликаних бактеріями родини Enterobacteriaceae, залежать від локалізації патологічного процесу. Вони не можуть бути диференційовані з іншими бактеріальними інфекціями на підставі одних лише симптомів і ознак захворювання, тому дуже важливо визначити збудника інфекції, дати видову і типову характеристику, чутливість до антибактеріальних засобів. Системне вивчення видових, типових і інших особливостей мікроорганізмів − збудників інфекцій − має істотне значення для з'ясування патогенезу інфекцій і визначення методів їхньої терапії.
Залежно від локалізації патологічного процесу для дослідження використовують виділення статевих органів, сечу, кров, спинномозкову рідину та ін. У пофарбованих мазках виявляють грамнегативні короткі палички, що можуть утворювати ланцюжки; вони дуже схожі один на одного і лише наявність широких капсул має діагностичне значення для роду Klebsiella. Досліджуваний матеріал одночасно наносять на кров'яний агар і середовища, що містять спеціальні
барвники і вуглеводи, що дозволяє швидко диференціювати ферганські і неферганські колонії. Бактерії, виділені на цих середовищах, надалі ідентифікують за допомогою біохімічних і серологічних реакцій. Можлива швидка попередня ідентифікація при можливості мікроорганізмів ферментувати лактозу.
Виражену протимікробну дію щодо бактерій родини Enterobacteriaceae має ампіцилін, тетрацикліни, аміноглікозиди, поліміксини, сульфаніламіди. Однак чутливість до цих препаратів в окремих штамів різна, тому її необхідно визначати лабораторними методами. Нерідко зустрічається множинна стійкість до ліків.
При лікуванні ефект може бути досягнутий при одночасному застосуванні аміноглікозидів і фурагіну чи ампіциліну; триметоприму із сульфаметоксазолом і поліміксином. Доцільні й інші комбінації антибактеріальних засобів, призначення препаратів, що потенціюють дію антибіотиків, що підвищують захисні механізми організму хворих, що сприяють доставці антибактеріальних засобів до вогнищ ураження й забезпечують їхню терапевтичну концентрацію в тканинах і сироватці крові.
Профілактика й ефективність боротьби з цими інфекціями багато в чому залежать від дотримання чистоти рук, суворого дотримання правил асептики й антисептики (стерилізації інструментарію, дезінфекції устаткування), обережності в призначенні внутрішньовенних препаратів і особистої гігієни сечостатевих шляхів. Дуже важливо пам'ятати, що багато грамнегативних бактерій є патогенними і викликають розвиток захворювання в ослабленому організмі. У лікувальних установах ці бактерії нерідко переносяться персоналом, а також через інструменти чи препарати для парентерального введення (2).
1. МОРФОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Мікроорганізми родини Enterobacteriaceae являють собою палички довжиною 1-5 мкм і товщиною 0,4-0,8 мкм. Вони добре фарбуються аніліновими барвниками, грамнегативні. Багато з них рухливі завдяки наявності перетрихіально розташованих джгутиків і можуть мати капсулу. Спор не утворюють. У багатьох штамів, які належать до різних родів ентеробактерій: ешерихіям, сальмонелам, шигелам, клебсієлам, протеям, виявлені ворсинки (пілі) загального типу, а у деяких штамів – статеві ворсинки (F-пілі), з якими зв’язані їх адгезивні та інші властивості (3,4).
Ешерихії
Рід E. coli представлений єдиним видом і багато чисельними біоварами і сероварами, які розрізняються деякими біохімічними, антигенними і патогенними властивостями.
Кишкову паличку (рис. 1.1) вперше виділив із фекалій людини в 1885 році німецький педіатр і мікробіолог Теодор Ешерих. В подальшому було встановлено, що подібні бактерії широко розповсюдженні в природі. Вони зустрічаються в кишечнику майже всіх видів ссавців, птахів, риб, а також в воді, ґрунті і інших об’єктах навколишнього середовища.
За своїми морфологічними і тинкторіальними властивостями E. coli нагадує інші ентеробактерії. Це дрібні грамнегативні палички довжиною 2-3 мкм, товщиною 0,5-0,7 мкм з закругленими кінцями, в мазках розташовуються не упорядковано. Серед кишкових паличок зустрічаються рухливі і не рухливі варіанти. Деякі штами мають виражену капсулу і утворюють слизові колонії. S-форми E. coli на поживному агарі формують опалово-мутнуваті, слабко випуклі, вологі, з рівними краями і блискучою поверхнею колонії, які легко суспендуються в сольовому розчині. Перехідні S- і R-форми утворюють пласкі, з шершавою поверхнею і нерівними краями колонії, які погано суспендуються в сольовому розчині. Кишкова паличка викликає рівномірне помутніння рідини поживного середовища з утворенням невеликого осаду (3).
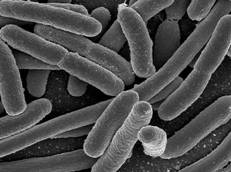
Рисунок 1.1 - Escherichia coli (за (5))
Сальмонели
До роду Salmonella (рис. 1.2) відносяться збудники черевного тифу, паратифу і сальмонельозів. Назва роду походить від імені американського дослідника Даніеля Сальмона. Сальмонели підрозділені на 4 підроду, які включають 65 серогруп і близько 2000 сероварів.
Сальмонели відповідають в основному тим ознакам, які присутні всім ентеробактеріям. При мікроскопії мазків, забарвлених за грамом, черевнотифозні і паратифозні бактерії мають вигляд паличок червоного кольору із заокругленими кінцями розміром 0,5-0,8×1-3 мкм, активно рухливі у висячій чи роздавленій краплі.
Ріст у МПБ супроводжується помутнінням. На МПА виростають ніжні, круглі, гладенькі, прозорі або напівпрозорі колонії розміром 2-4 мкм. Однак колонії тифозних мікробів, що мають Vi-антиген, каламутні. У S. Schottmuelleri колонії грубіші, через кілька днів по периферії колонії утворюється слизовий валик.
На середовищах Ендо, Левіна, Плоскирєва колонії безбарвні, прозорі, часом рожевуваті (Ендо) або злегка голубуваті (Левіна). На вісмут-сульфатному агарі черевнотифозні мікроби утворюють колонії чорного кольору, іноді зі світлим обідком. Паратифозні бактерії на цьому середовищі можуть утворювати коричневі або зеленуваті колонії. Після зняття колонії залишається чорний слід (3,6).
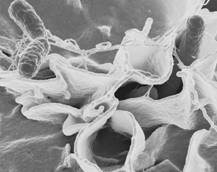
Рисунок 1.2 - Salmonella (за (7))
Шигели
Бактерії роду Shigella (рис. 1.3) є збудниками бактеріальної дизентерії або шигельозу.
Перші припущення про роль бактерій в етіології дизентерії були зроблені А. Шентемесом і Ф. Відалем в 1888 році.
Дизентерія – поліетіологічне захворювання. Його викликають різні види бактерій, які називаються шигелами. В теперішній час вони віднесені до роду Shigella, який підрозділяється на чотири групи: A,B,C і D – і більше двох десятків сероварів.
За своїми морфологічними властивостями шигел мало відрізняються від ешеріхій і сальмонел. Однак вони позбавленні джгутиків і тому є нерухомими бактеріями. Багато штамів шигел мають ворсинки загального типу і статеві ворсинки. Різні види шигел ідентичні за своїми морфологічними властивостями (3).
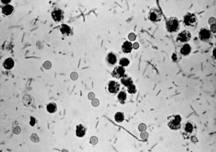
Рисунок 1.3 - Shigella (за (8))
Клебсієли
У мазках забарвлених за Грамом, капсульні форми клебсієл мають вигляд грамнегативних, еліпсоподібних, товстих паличок довжиною 5-8 мкм і шириною 3-5 мкм. Без капсульні форми мають менші розміри (0,3-0,6×1-3 мкм), поодиноке, парне або ланцюжкове розташування (6).
Протей
Усі види роду Proteus (рис. 1.4) являють собою грамнегативні прямі палички розмірами 0,4-0,6×1,0-3,0 мкм. Зустрічаються кокоподібні, нитковидні форми. Спор і капсул не утворюють, мають перетрихіально розташовані джгутики (3).
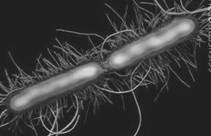
Рисунок 1.4 - P. vulgaris (за (9))
2. ФІЗІОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Усі ентеробактерії є факультативними анаеробами, добре ростуть на середовищах з м’ясним екстрактом. Вони володіють вираженою ферментативною активністю, яка пов’язана з утворенням багато чисельних сахаридолітичних, протеолітичних і інших ферментів, як конституційних, так і індуцибельних. Разом з тим відмінності які спостерігаються в перерахованих ознаках мають таксономічне значення і використовуються для родинної і видової диференціації і ідентифікації ентеробактерій. Деякі з цих ознак слугують підставою для визначення окремих біоварів серед одного і того ж виду бактерій.
До основних ознак, які вказують на належність ентеробактерій до одного з п’яти триб відносяться:
1) характер продукту, який утворюється при зброджуванні глюкози: кислоти, бутандіол, їх суміші;
2) реакція з метиловим червоним;
3) утворення ацетилметилкарбінолу і позитивна реакція Фогеса-Проскауера;
4) відновлення нітратів;
5) утворення уреази;
6) ріст в присутності KCN і деякі інші.
Головною біохімічною ознакою, яка слугує для визначення родової і видової належності ентеробактерій, є їх здатність ферментувати різні вуглеводи до утворення кислоти або газу, утворення індолу, сірководню, декарбоксилаз амінокислот (лізину, орнітину та ін.), утилізація цитрату, тартрату та інші (3).
Ешерихії
Це факультативний анаероб, добре росте на звичайних поживних середовищах при слабо лужній реакції середовища і оптимальній температурі 37°С. Ріст і розмноження бактерій можливі при доволі значних коливаннях рН середовища і температурного режиму.
Кишкова паличка має найбільш високу ферментативну активність. Вона утилізує ацетат в якості єдиного джерела вуглецю, відновлює нітрати до нітритів. Ферментує глюкозу та інші вуглеводи з утворенням пірувату, який потім перетворюється в молочну, оцтову, мурашину кислоти виділяються CO![]() і H
і H![]() . Більшість штамів ферментує лактозу. Однак зустрічаються штами, які повільно зброджують лактозу або зовсім не володіють цією здатністю (3).
. Більшість штамів ферментує лактозу. Однак зустрічаються штами, які повільно зброджують лактозу або зовсім не володіють цією здатністю (3).
Сальмонели
Вони є ферментативно активними бактеріями, хоча деякі поступаються в цьому відношенні ешеріхіям. Диференціація багато чисельних представників даного роду проводиться на основі їх ферментативної активності і антигенної структури наведено в таблиці 2.1 (3).
Таблиця 2.1 - Ферментативні властивості ешерихій і тифозно-паратифозних бактерій (за (6))
| Вид | Лактоза | Глюкоза | Мальтоза | Маніт | Сахароза | Індол | H |
Escherichia coli Salmonella typhi Salmonella parathyphi A Salmonella schottmuelleri | кг - - - | кг к кг кг | кг к кг кг | кг к кг кг | кг± - - - | + - - - | ± + - + |
Примітка: к-утворення кислоти; кг-утворення кислоти і газу; + - виявлення ознаки; − відсутність ознаки; ± - можливе виявлення ознак.
Шигели
Збудники дизентерії хемоорганогетеротрофні, невимогливі к поживним середовищам. На щільних середовищах при виділенні із організму хворого утворюються, як правило, S-форми колоній. Шигели виду sonnei утворюють два типи колоній − S- (I фаза) і R- (II фаза). Бактерії I фази при пересівах утворюють обидва типи колоній. Оптимальна температура росту 37°С, рН середовища 7,2.
Шигели менш ферментативно активні, ніж інші ентеробактерії: при зброджуванні глюкози та інших вуглеводів утворюються кислі продукти без газоутворення.
Шигели не розщеплюють лактозу і сахарозу, за виключенням S. sonnei, які повільно (на 2-у добу) розщеплюють ці сахариди, наведено в таблиці 2.2 (3).
Таблиця 2.2 - Біохімічні властивості шигел (за (6))
| Вид | Ферментація вуглеводів | Індол | Каталаза | |||||
| лактоза | глюкоза | мальтоза | маніт | дульцит | сахароза | |||
S. dysenteriae S. flexneri S. boydii S. sonnei | - - - + повільно | + + + + | - + ± + | - + + + | - + + - | - - - + повільно | - - + - | - - - + |
Клебсієли
Вони невимогливі до поживного субстрату. Добре ростуть на основних поживних середовищах, утворюючи куполоподібні блискучі слизові колонії. Викликають інтенсивне помутніння середовища при рості у бульйоні.
Більшість штамів ферментують глюкозу з утворенням кислоти і газу; використовують глюкозу і цитрат в якості єдиного джерела вуглецю, а аміак - як джерело азоту.
Диференціація різних видів клебсієл проводиться на основі їх різних здатностей ферментувати вуглеводи, утворювати уреазу і лізиндекарбоксилазу, утилізувати цитрат та інших властивостей наведено в таблиці 2.3.
Клебсієли є факультативними анаеробами. Оптимальна температура для їх росту 35-37°С (3).
Таблиця 2.3 - Диференціальні ознаки клебсієл (за (6))
| Вид | Ферментація | Індол | Реакція Фогеса-Проскауера | Утилізація цитрату | |||
| Глюкоза | Лактоза | Сахароза | Сечовина | ||||
K. pneumoniae K. oxytoca K. planticola K. terrigena K. ozaenae K.rhinoskleromatis | + + + + ± ± | + + + + ± − | + + + + + ± | + + + - - - | - + ± - - - | + + + + - - | + + + ± ± - |
Протеї
Вони не вимогливі до поживного субстрату, добре ростуть на основних поживних середовища. На середовищах Ендо і Плоскирєва виростають прозорі блискучі безбарвні колонії. На вісмут-сульфатному агарі через 48 годин утворюються сірувато-коричневі колонії, під якими формується чорно-коричнева зона. Н-форма (джгутикова) на простому агарі дає характерний повзучий ріст або „феномен роїння”. Культура має неприємний гнильний запах. При посіві у стовпчик напіврідкого агару визначають рухливість. Повзучий характер росту протею використовують для виділення чистих культур посівом у конденсаційну воду скошеного агару за методом Шукевича. Протеї є факультативними анаеробами. Ростуть при температурі 20-37°С.
Ферментують багато вуглеводів з утворенням кислих продуктів, глюкозу розщеплюють з утворенням кислот і невеликої кількості газу. Різні види відрізняються один від одного за ферментацією вуглеводів, утворенню індолу, уреази, сірководню, орнітиндекарбоксилази та іншим ознакам, наведено в таблиці 2.4 (3,6).
Таблиця 2.4 - Диференціальні ознаки бактерій роду Proteus (за (6))
| Вид | Ферментація | Утворення | Орнітин-декарбокси- лаза | Реакція Фогеса-Проскауера | ||||||
| мальтози | сахарози | ксилози | індолу | H | ||||||
P. vulgaris P. mirabilis P. myxofaciens P. penneri | + - + + | + - + + | + + - + | + - - - | + + - ± | - + - - | - ± + - | |||
3. МЕТАБОЛІЗМ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Більшість ентеробактерій можуть використовувати в якості субстратів для дихального метаболізму велику кількість простих органічних сполук – органічні кислоти, амінокислоти, вуглеводи. В аеробних умовах, всі ці бактерії добре ростуть на звичайних складних бактеріологічних середовищах, азотисті компоненти яких (амінокислоти і пептиди) забезпечують їх окислюючими субстратами. В анаеробних умовах, однак, ріст стає строго залежним від наявності зброджує мого вуглеводу. Деякі моносахариди, дисахариди і поліспирти зброджуються всіма представниками даної групи. Використання полісахаридів менш розповсюджено.
Хоча ентеробактерії вирощують на складних середовищах, мінімальні харчові потреби цих організмів, зазвичай прості. Представники багатьох родів взагалі не потребують факторів росту (наприклад Escherichia coli, багато видів Salmonella), а у ауксотрофних організмів ці потреби зазвичай прості. Особливо розповсюджена потреба в нікотиновій кислоті, що характерна для багатьох видів роду Proteus. У Salmonella typhi є специфічна потреба в триптофані.
Дослідження механізму регуляції біосинтезу амінокислот у багатьох ентеробактерій виявило характерні особливості цього процесу, які відрізняють данні організми від всіх інших бактерій. Наприклад, перша стадія в біосинтезі амінокислот родини аспартату, перетворення аспарагінової кислоти в аспартилфосфат, у ентеробактерій завжди здійснюється трьома ізофункціональними аспартаткіназами: аспартаткіназою I, яка і інгібується, і репресується треоніном, аспартаткіназою II, яка репресується, але не інгібується метіоніном, і аспартаткіназою III, яка і інгібується, і репресується лізином. Цей тип регуляції аспарагінового шляху не був виявлений ні в однієї бактерії не кишкової групи.
ГЦ-склад ДНК ентеробактерій варіює в широких межах – від 37 до 63%. За виключенням роду Proteus, представники якого суттєво відрізняються за нуклеотидним складом. ГЦ-склад ДНК споріднених організмів трьох видів - Salmonella, Escherichia і Shigella – розрізняються зовсім мало. Діапазон змінення
ГЦ-складу для „класичних” бактерій родини Enterobacteriaceae (від 37 до 59%) дуже близький до діапазону для представників родини з полярним джгутикуванням (від 39 до 63%) (10).
Бродіння
Зброджування сахаридів у ентеробактерій відбувається за шляхом Ембдена-Мейєргофа. Продукти бродіння у різних представників групи розрізняються і якісно, і кількісно. Однак у цих процесів є одна характерна біохімічна властивість, яка рідко зустрічається при інших типах бактеріального бродіння. Мова йде про особливий спосіб розщеплення однієї з проміжних сполук – піровиноградної кислоти (ПВК), що веде до утворення мурашиної кислоти:
СН3СОСООН + КоАSН→СН3СОSКоА+ НСООН.
Таким чином, мурашина кислота часто є основним кінцевим продуктом бродіння. Вона накопичується, хоча не завжди, оскільки деяки з цих бактерій володіють формнатгідрогенліазою, яка розщеплює мурашину кислоту до СО2 і Н2:
НСООН → СО2+Н2.
У таких організмів замість мурашиної кислоти як кінцевого продукту бродіння утворюються еквімолярні кількості Н2 і СО2.
Найбільш розповсюдженим типом перетворення сахаридів при бродінні у ентеробактерій є так зване бродіння змішаного типу, яке приводить до утворення молочної, оцтової, бурштинової і мурашиної кислот (або СО2 і Н2), а також етилового спирту. Такий тип бродіння властив представникам родів Salmonella, Escherichia, Shigella і Proteus. Відношення кількості різних кінцевих продуктів різниться як від штаму до штаму, так і для одного штаму при рості в різних умовах, наприклад при різних значеннях рН. Така різниця відображає ту обставину, що кінцеві продукти утворюються з піровиноградної кислоти трьома незалежними шляхами (рис. 3.1). Утворення газу в результаті зброджування сахаридів є дуже важливою ознакою для ентеробактерій – газоутворювальні бактерії роду Escherichia відрізняються за цим критерієм від патогенних видів групи Shigella і Salmonella typhi, які зброджують сахариди без виділення газу. В випадку простого бродіння змішаного типу газ може утворюватися тільки за рахунок розщеплення мурашиної кислоти; відповідно, утворення газу відображає наявність форміатгідрогенліази. Цей ферментний комплекс, звісно, не суттєвий для бродіння і може загубитися в результаті мутації без зміни здатності бактерії до бродіння. І дійсно, дослід показує, що в природі існують „неаерогенні”, тобто не утворюючі газоподібних продуктів штами такого типового продуценту їх, як Escherichia coli. Тому, хоча виділення газу являє собою корисну ознаку при класифікації ентеробактерій, даний критерій не є безпомилковим.
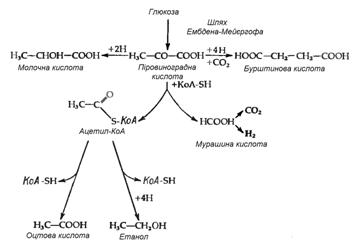
Рисунок 3.1 - Шляхи утворення із ПВК типових кінцевих продуктів кислотного бродіння змішаного типу (за (10))
Іншою ознакою, яка має велике значення для ідентифікації ентеробактерій, є здатність зброджувати дисахарид лактозу, що обумовлено наявністю у цих організмів β-галактозідази. Ефективність зброджування лактози залежить ще і від наявності специфічної пермеази галактозидів, яка сприяє проникненю лактози в клітину. Штами які мають β-галактозідазу, але позбавленні пермеази, не можуть поглинати лактозу зі швидкістю, яка достатня для інтенсивного бродіння, і зазвичай класифікуються як не здатні до зародження цього сахариду. Зброджування лактози властиво Escherichia, але відсутні у Salmonella, Shigella і Proteus. Деякі штами Shigella утворюють β-галактозідазу, але не можуть зброджувати лактозу із-за відсутності пермеази (10).
4. ГЕНЕТИКА БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Відкриття явища переносу генів у ентеробактерій при кон’югації і трансдукції дозволило доволі детально дослідити генетичні родинні відносини між деякими її представниками. Утворення хромосомних гібридів між E. coli і бактеріями родів Salmonella і Shigella каже про дуже високу ступінь генетичної гомології цих бактерій, що підтверджується і порівнянням хромосомних карт двох найбільш вивчених в цьому відношенні видів − Salmonella typhimurium і Escherichia coli. Як видно на рис. 4.1 і 4.2, де представленні в спрощеному вигляді хромосомні карти цих двох організмів, багато маркерів розташовуються на обох хромосомах в однакових локусах.
Про близьку генетичну спорідненість груп Escherichia, Salmonella і Shigella свідчить також висока ступень ДНК-ДНК-гібридизації in vitro. Утворення ж хромосомних гібридів між бактеріями цієї підгрупи і представниками інших родів ентеробактерій (Proteus, Enterobacter) відбувається дуже рідко. Крім того, випробування по гібридизації ДНК-ДНК вказують, що ступень генетичної гомології між представниками підгрупи Escherichia− Salmonella− Shigella та іншими групами ентеробактерій доволі низька. Ентеробактерії, що належать до різних родів, можуть отримувати плазміни від донорних штамів E. coli при кон’югації і потім зберігати їх як позахромосомні елементи (ефективність кон’югації варіює в широких межах; таблиця 4.1). Таким чином серед бактерій кишкової групи можуть розповсюджуватися F-фактори, які містять додаткові гени (наприклад F-lac), і R-фактори, які визначають стійкість до різноманітних лікарських засобів (10).
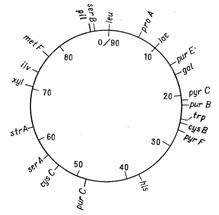
Рисунок 4.1 - Генетична карта E. coli К12 в спрощеному вигляді (за (10))
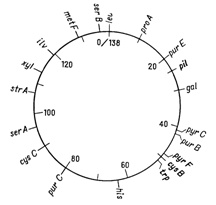
Рисунок 4.2 - Генетична карта S. typhimurium в спрощеному вигляді (за (10))
5. ЕКОЛОГІЯ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Середовищем проживання для більшості ентеробактерій є кишковий тракт хребетних тварин і людини.
В організмі людини багато ентеробактерій містяться в складі мікробних біоценозів тонкого і товстого кишечника. Патогенні види зустрічаються тільки у хворих і бактеріоносіїв.
З випорожненнями людей і тварин ентеробактерії потрапляють в навколишнє середовище. В ній вони можуть зберігатися на протязі різних строків в залежності від виду і умов. Деякі з них (E. coli) використовуються в санітарній мікробіології в якості показників фекального забруднення навколишнього середовища (3).
Кишкова паличка наприклад може зберігатися в воді і ґрунті декілька місяців (11).
Широке розповсюдження сальмонел в природі і різноманіття шляхів, якими вони проникають в організм людини, пояснюється їх генетичною пластичністю. Сальмонели легко знаходять екологічні ніші і адаптуються до найрізноманітніших умов (12).
Клебсієли – широко розповсюджені бактерії. Вони порівняно стійкі до факторів навколишнього середовища завдяки наявності капсули і можуть на протязі тривалого часу зберігатися в ґрунті, воді, приміщеннях. Разом з тим вони зустрічаються в складі мікробних біоценозів в організмі людини і тварин (3).
Протеї P. vulgaris і P. mirabilis є мешканцями кишечника багатьох тварин, виявляються також в стічних водах і ґрунті. Всі види протеїв можуть виділятися з хворих людей (3).
6. АНТИГЕНИ БАКТЕРІЙ РОДИНИ ENTEROBACTERIACEAE
Антигенна будова служить одним із суттєвих критеріїв, на яких основана класифікація, а також ідентифікація ентеробактерії. Розрізняють три основних типи антигенів: 1) О-соматичний антиген; 2) Н-джгутиковий антиген; 3)К-антигени. О-антиген є складовою частиною ліпополісахариду (ЛПС) зовнішнього шару клітинної стінки. Специфічність О-антигену визначається детермінантними сахаридами (гексозами і аміносахаридами), ковалентно зв’язаними з базисною частиною ЛПС. Н-антиген локалізується в джгутиках клітини. Він складається з білка флагеліну. Капсульні антигени К-антигени, так як і О-антигени, містяться в ЛПС клітинної стінки, але в більш поверхневому шарі. Вони маскують О-антигени.
В зв’язку з цим для визначення О-антигену у бактерій, які володіють К-антигеном, необхідно зруйнувати останній шляхом кип’ятіння або автоклавування культури. К-антигени за хімічними властивостями відносяться до кислих полісахаридів. До К-антигенів відносяться К-антигени E. coli, М-антигени Salmonella schottmuelleri, Vi-антиген збудника черевного тифу, який виявляється також у S. parathyphi C та деяких штамів E. coli. Всі ці антигени характеризуються імуннохімічною специфічністю, що дозволяє диференціювати роди і види, а також виділяти серед них серогрупи і серологічні варіанти (серовари). Антигенну будову е