Химические процессы в биологическом организме
Содержание
1. Опишите пути внедрения ферментативных методов синтеза в химическое производство
2. Найдите способ определения содержания аминокислот триптофана и цистеина в составе белков. Напишите соответствующие уравнения реакции
3. Покажите специфику строения и состава структурных белков биологической мембраны
4. Дайте сравнительную характеристику всех видов РНК. Оформите ответ в виде таблицы, указав молярную массу, минорные основания, углеводы, структуру, место локализации, функции
5. Фермент лактатдегидрогенеза окисляет молочную кислоту в пировиноградную. Покажите с помощью уравнения данной реакции механизм действия кофермента НАД
6. Объясните комплементарный механизм репликации ДНК и его роль в обеспечении специфичности воспроизведения структуры. Дайте соответствующие схемы
7. Какие метаболиты образуются в результате окислительного дезаминирования глутаминовой кислоты, аспарагиновой кислоты и аланина? Напишите уравнения реакций дальнейшего превращения метаболитов в организме. Назовите ферменты, катализирующие эти процессы
8. Напишите уравнение реакции фосфолирирования галактозы при участии соответствующей киназы и дальнейшего перехода фасфарного эфира галактозы во фруктозу 6-фосфат. Дайте полное название метаболитов и ферментов, ускоряющих эти реакции
9. HS-KoA необходим для активирования высших жирных кислот. Укажите пути его высвобождения в организме животных и напишите соответствующие уравнения реакций
10. Охарактеризуйте окислительное фосфорилирование на примере окислительного декарбоксилирования пировиноградной кислоты. Чем отличается окислительное фосфорилирование от фотосинтетического?
11. Охарактеризуйте сходство и различие в действии соматотропина (гормона роста) и инсулина. Укажите, к какому классу относится каждый из них
12. На примере превращений 3-фофоглицеринового альдегида покажите взаимосвязь углеводного и липидного обменов. Напишите уравнения реакций
Используемая литература
1. Опишите пути внедрения ферментативных методов синтеза в химическое производство
Ферменты, или энзимы, представляют собой высокоспециализированный класс веществ белковой природы, используемый живыми организмами для осуществления с высокой скоростью многих тысяч взаимосвязанных химических реакций, включая синтез, распад и взаимопревращение огромного множества разнообразных химических соединений. Жизнь и многообразие ее проявлений – сложная совокупность химических реакций, катализируемых специфическими ферментами. И.П. Павлов считал ферменты «возбудителями всех химических превращений» у живых существ. Как известно, важнейшим свойством живого организма является обмен веществ, ускоряющим аппаратом, основой молекулярных механизмов, интенсивности которого являются ферменты. «Вся тайна животной жизни,– писал Д.И. Менделеев,– заключается в непрерывных химических превращениях веществ, входящих в состав животных тканей».
В настоящее время теоретические и практические достижения энзимологии используются в решении многих проблем биохимии и молекулярной биологии, включая их сравнительное и эволюционное рассмотрение. «Под знаком молекулярной энзимологии,– говорил на III Всесоюзном биохимическом съезде (1974) А.Е. Браунштейн,– развивается и встречное течение – реконструкция или интеграция, восходящая от молекулярного яруса к высшим уровням структурно-функциональной организации живого и пронизывающая весь комплекс актуальных проблем биологии и медицины».
Ферменты обеспечивают осуществление таких важнейших процессов жизнедеятельности, как экспрессия (реализация) наследственной информации, биоэнергетика, синтез и распад биомолекул (обмен веществ). Изучение их способствует проникновению в суть и сокровенные тайны того загадочного явления, которое мы называем жизнью. Этими обстоятельствами может быть объяснено пристальное внимание исследователей к проблемам структуры, функций и молекулярных механизмов действия ферментов.
От неорганических катализаторов ферменты отличаются рядом характерных особенностей. Прежде всего, ферменты чрезвычайно эффективны и проявляют в миллионы и миллиарды раз более высокую каталитическую активность в условиях умеренной температуры (температура тела), нормального давления и в области близких к нейтральным значениям рН среды.
Полипептидная цепь способна самопроизвольно формировать и удерживать особую пространственную структуру. Исходя из формы белковых молекул белки делят на фибриллярные и глобулярные. В глобулярных белках одна или несколько полипептидных цепей свернуты в компактную структуру сферической формы, или глобулу. Обычно эти белки хорошо растворимы в воде. К их числу относятся почти все ферменты, транспортные белки крови и многие запасные белки. Фибриллярные белки представляют собой нитевидные молекулы, скрепленные друг с другом поперечными связями и образующие длинные волокна или слоистые структуры. Они обладают высокой механической прочностью, нерастворимы в воде и выполняют главным образом структурные и защитные функции. Типичными представителями таких белков являются кератины волос и шерсти, фиброин шелка, коллаген сухожилий.
Порядок расположения ковалентно связанных аминокислот в полипептидной цепи называют аминокислотной последовательностью, или первичной структурой белков. Первичная структура каждого белка, кодируемая соответствующим геном, постоянна и несет в себе всю информацию, необходимую для формирования структур более высокого уровня. Потенциально возможное число белков, которые могут образоваться из 20 аминокислот, практически не ограничено.
В результате взаимодействия боковых групп аминокислотных остатков отдельные относительно небольшие участки полипептидной цепи принимают ту или иную конформацию (тип укладки), известную как вторичная структура белков. Наиболее характерными элементами ее являются периодически повторяющиеся a-спираль и b-структура. Вторичная структура весьма стабильна. Так как она в значительной мере определяется аминокислотной последовательностью соответствующего участка белка, становится возможным ее предсказание с определенной степенью вероятности. Термин «a -спираль» был введен американским биохимиком Л. Полингом, описавшим укладку полипептидной цепи в белке a -кератине в виде правосторонней спирали (a -спираль можно сравнить со шнуром от телефонной трубки). На каждый виток такой спирали в белке приходится 3,6 аминокислотных остатков. Это означает, что группа —С= О одной пептидной связи образует водородную связь с группой —NH другой пептидной связи, отстоящей от первой на четыре аминокислотных остатка. В среднем каждый a -спиральный участок включает до 15 аминокислот, что соответствует 3-4 оборотам спирали. Но в каждом отдельном белке длина спирали может сильно отличаться от этой величины. В поперечном сечении a -спираль имеет вид диска, от которого наружу направлены боковые цепи аминокислот.
b-структура, или b-складчатый слой, может быть образована несколькими участками полипептидной цепи. Эти участки растянуты и уложены параллельно друг другу, связываясь между собой водородными связями, которые возникают между пептидными связями. Они могут быть ориентированы в одном и том же или в противоположных направлениях (направление движения вдоль полипептидной цепи принято считать от N-конца к С-концу). В первом случае складчатый слой называют параллельным, во втором — антипараллельным. Последний образуется, когда пептидная цепь делает резкий поворот вспять, образуя изгиб (b-изгиб). Боковые цепи аминокислот ориентированы перпендикулярно плоскости b-слоя.
Относительное содержание a -спиральных участков и b-структур может широко варьироваться в разных белках. Существуют белки с преобладанием a-спиралей (около 75% аминокислот в миоглобине и гемоглобине), а основным типом укладки цепи во многих фибриллярных белках (в том числе фиброин шелка, b-кератин) является b-структура. Участки полипептидной цепи, которые нельзя отнести ни к одной из вышеописанных конформаций, называют соединительными петлями. Их структура определяется главным образом взаимодействиями между боковыми цепями аминокислот, и в молекуле любого белка она укладывается строго определенным образом.
Третичной структурой называют пространственное строение глобулярных белков. Но часто это понятие относят к характерному для каждого конкретного белка способу сворачивания полипептидной цепи в пространстве. Третичная структура формируется полипептидной цепью белка самопроизвольно, по-видимому, по определенному пути (путям) свертывания с предварительным образованием элементов вторичной структуры. Если стабильность вторичной структуры обусловлена водородными связями, то третичная структура фиксируется разнообразной системой нековалентных взаимодействий: водородными, ионными, межмолекулярными взаимодействиями, а также гидрофобными контактами между боковыми цепями неполярных аминокислотных остатков. В некоторых белках третичная структура дополнительно стабилизируется за счет образования дисульфидных связей (—S—S—-связей) между остатками цистеина. Как правило, внутри белковой глобулы расположены боковые цепи гидрофобных аминокислот, собранные в ядро (их перенос внутрь глобулы белка выгоден термодинамически), а на периферии находятся гидрофильные остатки и часть гидрофобных. Белковую глобулу окружает несколько сотен молекул гидратной воды, необходимой для стабильности молекулы белка и нередко участвующей в его функционировании. Третичная структура подвижна, отдельные ее участки могут смещаться, что приводит к конформационным переходам, которые играют значительную роль во взаимодействии белка с другими молекулами. Третичная структура является основой функциональных свойств белка. Она определяет образование в белке ансамблей функциональных групп — активных центров и зон связывания, придает им необходимую геометрию, позволяет создать внутреннюю среду, являющуюся предпосылкой протекания многих реакций, обеспечивает взаимодействие с другими белками.
Третичная структура белков однозначно соответствует его первичной структуре; вероятно, существует еще нерасшифрованный стереохимический код, определяющий характер свертывания белка. Однако один и тот же способ укладки в пространстве обычно соответствует не единственной первичной структуре, а целому семейству структур, в которых совпадать может лишь небольшая доля (до 20-30%) аминокислотных остатков, но при этом в определенных местах цепи сходство аминокислотных остатков сохраняется. Результатом является образование обширных семейств белков, характеризующихся близкой третичной и более или менее сходной первичной структурой и, как правило, общностью функции. Таковы, например, белки организмов разных видов, несущие одинаковую функцию и эволюционно родственные: миоглобины и гемоглобины, трипсин, химотрипсин, эластаза и другие протеиназы животных.
Нередко, особенно в крупных белках, сворачивание полипептидной цепи проходит через формирование отдельными участками цепи более или менее автономных элементов пространственной структуры — доменов, которые могут обладать функциональной автономией, будучи ответственными за ту или иную биологическую активность белка. Так, N-концевые домены белков системы свертывания крови обеспечивают их присоединение к клеточной мембране.
Существует много белков, молекулы которых представляют собой ансамбль из глобул (субъединиц), удерживаемых вместе за счет гидрофобных взаимодействий, водородных или ионных связей. Такие комплексы называют олигомерными, мультимерными или субъединичными белками. Укладку субъединиц в функционально активном белковом комплексе называют четвертичной структурой белка. Некоторые белки способны образовывать структуры более высоких порядков, например, полиферментные комплексы, протяженные структуры (белки оболочек бактериофагов), надмолекулярные комплексы, функционирующие как единое целое (например, рибосомы или компоненты дыхательной цепи митохондрий). Четвертичная структура позволяет создать молекулы необычной геометрии. Так, у ферритина, образованного 24 субъединицами, имеется внутренняя полость, благодаря которой белку удается связать до 3000 ионов железа. Кроме того, четвертичная структура позволяет в одной молекуле выполнять несколько различных функций. В триптофансинтетазе совмещены ферменты, ответственные за несколько последовательных стадий синтеза аминокислоты триптофана.
3. Покажите специфику строения и состава структурных белков биологической мембраны
Первичная структура белков определяет все остальные уровни организации белковой молекулы. Поэтому при изучении биологической функции различных белков важно знание этой структуры. Первым белком, для которого была установлена аминокислотная последовательность, был гормон поджелудочной железы — инсулин. Эта работа, потребовавшая 11 лет, была выполнена английским биохимиком Ф. Сенгером (1954). Он определил расположение 51 аминокислоты в молекуле гормона и показал, что она состоит из 2-х цепей, соединенных дисульфидными связями. Позже большая часть работ по установлению первичной структуры белков была автоматизирована. С развитием методов генетической инженерии появилась возможность еще более ускорить этот процесс, определяя первичную структуру белков в соответствии с результатами анализа нуклеотидной последовательности в генах, кодирующих эти белки. Вторичную и третичную структуру белков исследуют с помощью достаточно сложных физических методов, например, кругового дихроизма или рентгеноструктурного анализа белковых кристаллов. Третичная структура была впервые установлена английским биохимиком Дж. К. Кендрю (1957) для белка мышц — миоглобина.
синтез белок аминокислота
О количестве РНК нет точных данных, поскольку содержание ее в разных клетках в значительной степени определяется интенсивностью синтеза белка. На долю РНК приходится около 5–10% от общей массы клетки. Современная классификация различных типов клеточной РНК основывается на данных топографии, функции и молекулярной массы. Выделяют три главных вида РНК: матричную (информационную) – мРНК, которая составляет 2–3% от всей клеточной РНК; рибосомную – рРНК, составляющую 80–85% и транспортную – тРНК, которой около 16%. Эти 3 вида различаются нуклеотидным составом и функциями (табл. 1).
Таблица 1.
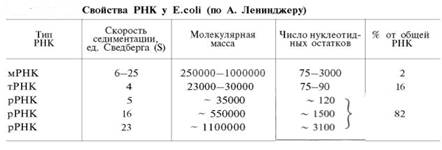
Окисление пирувата до ацетил-КоА происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название «пируватдегидрогеназный комплекс».
На I стадии этого процесса пируват теряет свою карбоксильную группу в результате взаимодействия с тиаминпирофосфатом (ТПФ) в составе активного центра фермента пируватдегидрогеназы (E1). На II стадии оксиэтильная группа комплекса E1–ТПФ–СНОН–СН3 окисляется с образованием ацетильной группы, которая одновременно переносится на амид липоевой кислоты (кофермент), связанной с ферментом дигидроли-поилацетилтрансферазой (Е2). Этот фермент катализирует III стадию – перенос ацетильной группы на коэнзим КоА (HS-KoA) с образованием конечного продукта ацетил-КоА, который является высокоэнергетическим (макроэргическим) соединением.
На IV стадии регенерируется окисленная форма липоамида из восстановленного комплекса дигидролипоамид–Е2. При участии фермента дигидролипоилдегидрогеназы (Е3) осуществляется перенос атомов водорода от восстановленных сульфгидрильных групп дигидролипоамида на ФАД, который выполняет роль простетической группы данного фермента и прочно с ним связан. На V стадии восстановленный ФАДН2 дигидро-липоилдегидрогеназы передает водород на кофермент НАД с образованием НАДН + Н+.
Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, ди-гидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 кофер-ментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E1, ли-поамид-Е2 и ФАД-Е3), а два – легко диссоциируют (HS-KoA и НАД).
Нуклеиновые кислоты (ДНК и РНК) относятся к сложным высокомолекулярным соединениям, состоят из небольшого числа индивидуальных химических компонентов более простого строения. Так, при полном гидролизе нуклеиновых кислот (нагревание в присутствии хлорной кислоты) в гидролизате обнаруживают пуриновые и пиримидиновые основания, углеводы (рибоза и дезоксирибоза) и фосфорную кислоту :
В молекуле ДНК углевод представлен дезоксирибозой, а в молекуле РНК – рибозой, отсюда их названия: дезоксирибонуклеиновая (ДНК) и рибонуклеиновая (РНК) кислоты. Кроме того, они содержат фосфорную кислоту, по два пуриновых и по два пиримидиновых основания; различия только в пиримидиновых основаниях: в ДНК содержится тимин, а в РНК – урацил. В составе ДНК и РНК открыты так называемые минорные (экзотические) азотистые основания (строение некоторых из них приводится далее).
Углеводы (рибоза и дезоксирибоза) в молекулах ДНК и РНК находятся в β-D-рибофуранозной форме:
В составе некоторых фаговых ДНК обнаружена молекула глюкозы, которая соединяется гликозидной связью с 5-оксиметилцитозином.
Основу структуры пуриновых и пиримидиновых оснований составляют два ароматических гетероциклических соединения – пиримидин и пурин :
Молекула пурина состоит из двух конденсированных колец: пиримидина и имидазола.
В составе нуклеиновых кислот встречаются три главных пиримидиновых основания: цитозин, урацил и тимин.
Активация фосфорилазы, как видно из рассмотренного выше цикла, должна сопровождаться снижением активности гликогенсинтетазы. Этот фермент также существует в двух формах, однако, в этом случае активная форма не фосфорилирована. При фосфорилировании соответствующей протеинкиназой она превращается в неактивную гликоген синтетазу.
Следовательно, изображенный выше цикл синтеза и фосфоролиза гликогена следует дополнить довольно сложной схемой взаимопревращений активных и неактивных форм ферментов, катализирующих эти процессы:
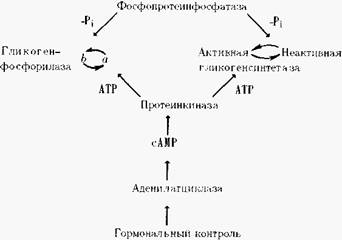
Необходимо иметь в виду, что и эта схема сильно упрощена. Выяснилось, что активность протеинкиназы может регулироваться калъмодулином - белком, связывающим ионы кальция, — и, следовательно, зависит от концентрации в клетке и этого вторичного мессенджера.
Аналогично, фосфорилированием и дефосфорилированием контролируется активность целого ряда других метаболических ферментов, в частности синтетазы жирных кислот, пируватдегидрогеназы. Важно подчеркнуть, что реакции фосфорилирования и дефосфорилирования коренным образом отличаются от аллостерической регуляции ферментов, поскольку они не находятся в прямой зависимости от биохимического статуса клетки и, следовательно, могут использоваться для изменения активности под воздействием внеклеточных сигналов, они удобны для регуляции на уровне организма. Известны и другие протеинкиназы, фосфорили-рующие остатки серина или треонина в белках, в том числе независимые от сАМР или контролируемые cGMP, диацилглицерином.
Выбор остатка серина, подвергаемого фосфорилированию, может зависеть от последовательности аминокислот, которые его окружают, Так, сАМР-зависимая протеинкиназа быка фосфорилирует гидроксильную группу серина в последовательности Arg-Arg-Ala-Ser-V al (Leu), причем соседствующие остатки аргинина, видимо, направляют фермент.
Фосфорилирование по остаткам тирозина, которое катализируется специфическими протеинкиназами (тирозинкиназами), встречается значительно реже, чем фосфорилирование остатков серина и треонина, — на долю фосфотирозина приходится около 0,1% всего связанного с белком фосфата. Однако биологическая роль этой реакции очень велика, так как она активно используется для передачи сигналов в биологических системах.
На первом этапе аденозилметионин (AdoMet), обычно являющийся донором метильной группы, выступает при алкилировании имидазольного кольца как донор фермента
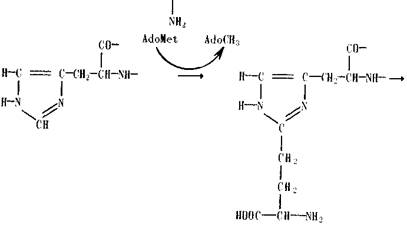
Затем происходит исчерпывающее метилирование аминогруппы присоединившегося фрагмента. В этом случае аденозилметионин является источником метильных групп, превращаясь в аденозилгомоцистеин (AdoHcy).
9. HS-KoA необходим для активирования высших жирных кислот. Укажите пути его высвобождения в организме животных и напишите соответствующие уравнения реакций
Получение чистых химических индивидуальных белков включает в себя как удаление небелковых примесей, так и разделение между собой собственно белковых компонентов. Последняя часть задачи в силу сходства физико-химических свойств белков особенно сложна, поэтому именно ее решение определяет выбор тех или иных схем выделения белка. При этом необходимо учитывать некоторые особенности поведения, присущие всем белкам.
К ним относится, прежде всего, способность белков подвергаться денатурации, т.е. претерпевать такие изменения пространственного строения, которые приводят к утрате или частичной потере функциональных свойств. Правда, денатурация во многих случаях обратима, однако эта обратимость не обеспечивается автоматически, а требует в каждом отдельном случае подбора специальных приемов. В то же время полностью или даже частично денатурированные белки весьма уязвимы для необратимых повреждений, в особенности для действия протеолитических ферментов, поэтому условий, способствующих денатурации, следует всемерно избегать.
Для предотвращения тепловой денатурации выделение белка проводят при низкой температуре, обычно при 4 °С. Необходимо также избегать крайних значений рН. Белки легко денатурируют при низких рН из-за протонирования отрицательно заряженных в нормальных условиях карбоксильных групп и возникающего вследствие этого резкого преобладания положительных зарядов, которое благоприятствует развертыванию компактной структуры. В щелочной среде при рН 10 и выше утрачиваются положительные заряды, обусловленные протонированием e-аминогрупп лизина, и опять-таки наступает декомпенсация зарядов на поверхности, дестабилизирующая глобулу.
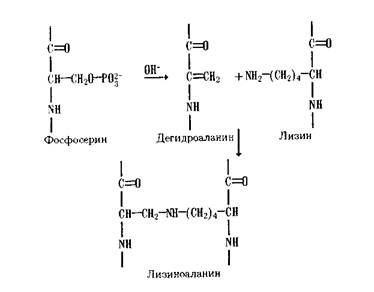
Фосфоролиз (от фосфор и греч. lysis - разрушение), ферментативная реакция расщепления химических связей в некоторых биологически важных соединениях с участием фосфорной кислоты; сопровождается включением фосфорильной группы (-Н2РО3) в образующиеся продукты. Ферменты, катализирующие реакцию, называются фосфорилазами. Они широко распространены в процессах обмена веществ у животных, растений и микроорганизмов. Фосфоролитическому расщеплению под действием ферментов могут подвергаться гликозидные (в гликогене), тиоэфирные (в ферментсубстратном комплексе, образующемся при окислении 3-фосфоглицеринового альдегида), углерод-углеродные (в ксилулозо-5-фосфате, в пировиноградной коферменте), фосфодиэфир-ные (в нуклеиновых к-тах) и углерод-азотные (в цитруллине) связи. Фосфоролиз играет важную роль в энергетике живых систем, т. к. фосфорильная группа, включённая в продукты реакции, под действием различных ферментов, в конечном счете, переносится на аденозиндифосфорную кислоту с образованием аденозинтрифосфорной кислоты (АТФ) - основного энергетического ресурса клеток.
В целом, реакция будет выглядеть следующим образом:
С6Н5СН = СН2 + 2РСl5 -> С6Н5СНС1 - СН2РСl4 • РСl5
Фосфорилирование по остаткам тирозина, которое катализируется специфическими протеинкиназами (тирозинкиназами), встречается значительно реже, чем фосфорилирование остатков серина и треонина - на долю фосфотирозина приходится около 0,1% всего связанного с белком фосфата. Однако биологическая роль этой реакции очень велика, так как она активно используется для передачи сигналов в биологических системах.
Так, инсулин, связываясь с рецептором - трансмембранным белком, вызывает активацию его домена, направленного в цитоплазму и являющегося тирозинкиназой. Тирозинкиназную активность индуцирует ряд факторов роста - белков, запускающих деление клеток по механизму, аналогичному описанному для инсулина. Рецепторы этих факторов, крупные трансмембранные белки - связывают их на наружной поверхности мембраны. Образование комплекса фактор рецептор передает сигнал через трансмембранную часть рецептора на его цитоплазматический домен, являющийся тирозинкиназой. Активированная таким образом тирозинкиназа модифицирует внутриклеточные белки, передавая сигнал далее.
Тирозинкиназы фосфорилируют остатки тирозина в некоторых внутриклеточных белках, вовлеченных в сложную систему регуляции клеточного деления, в которой также участвует и фосфорилирование белков по остаткам серина и треонина. Нарушение этой весьма сложной регуляторной сети может привести к неконтролируемому делению клеток, опухолевому росту. Характерно, что онкобелки, структурно близкие нормальным белкам животной клетки, но вызывающие ее трансформацию, превращение в опухолевую, нередко оказываются тирозин-киназами.
По завершении биосинтеза полипептидной цепи С-концевой участок такого белка, содержащий от 17 до 31 остатка преимущественно гидрофобных аминокислот, отщепляется, заменяясь следующей структурой:
Белок—СО—NH—СН 2СН2O—гликан—глюкозамин—фосфоинозит—диацилглицерин Этаноламин.
Гликолипидный фрагмент несколько различен у разных белков, отличаясь строением гликана, содержащего галактозу, набором жирных кислот в диацилглицериновом звене, среди которых отмечено присутствие пальмитиновой и миристиновой кислот.
Весь гликолипидный фрагмент удается в некоторых случаях удалить при действии протеиназы. Связь между глюкозамином и фосфоинозитом расщепляется при действии азотистой кислоты. Особенно характерно отщепление липида при действии фосфолипаз D или С, которые гидролизуют соответственно фосфодиэфирные связи между инозитом и фосфатидовой кислотой или фосфоинозитом и диацилглицерином, что приводит к отделению белка от мембраны. Имеются сведения, что фосфолипаза С каким-то образом участвует в механизме действия инсулина, отщепляя гликолипидный фрагмент от липопротеинлипазы.
1. Степанов В.М. Молекулярная химия. – М.: Высшая школа, 2002. – 329 с.
2. Ткачук А.Ф. Биохимия человека. – М.: Высшая школа, 2001. – 318 с.
3. Шатилов В. Р. Успехи биологической химии. – М.: Высшая школа, 1999. – 286 с.