Сальмонеллез у сельскохозяйственных животных и птиц
Краткая историческая справка
Характеристика возбудителя
Клинические признаки заболевания
Инкубационный период и патогенез
Сальмонеллез поросят
Сальмонеллез жеребят
Сальмонеллез собак
Сальмонеллез водоплавающих птиц
Сальмонеллезный (паратифозный) аборт лошадей
Сальмонеллезный (паратифозный) аборт овец
Эпизоотология
Лечение
Иммунитет
Профилактика и меры борьбы
Ветеринарно-санитарная экспертиза
Заключение
Список использованной литературы
Введение
Сальмонеллезы (Salmonellosis) - группа бактериальных болезней, преимущественно молодняка сельскохозяйственных и промысловых животных, характеризующихся при остром течении лихорадкой, явлениями септицемии, токсикоза и поражением кишечника, а при хроническом - воспалением легких. У взрослых животных (коров, кобыл, овец) болезнь проявляется абортами, а у людей - в виде пищевых токсикоинфекции.
Сальмонеллез вызывается паратифозными бактериями, относящимися к роду Salmonella. В настоящее время насчитывают свыше 1500 родственных разновидностей сальмонелл.
Возбудителями сальмонеллеза телят являются S. dublin, овец - S. abortus ovis, S. typhimurium, свиней - S. cholerae, лошадей - S. abortus equi, птиц - S. pullorum и S. galinarrum.
К сальмонеллезу восприимчивы все виды сельскохозяйственных животных и птиц. Болезнь поражает преимущественно молодняк; у взрослых животных сальмонеллез чаще протекает без заметных клинических признаков. Источником заражения являются взрослые животные - сальмонеллоносители, а также больной и переболевший сальмонеллезом молодняк. Заражение происходит через желудочно-кишечный тракт.
У животных нередко встречаются вторичные сальмонеллезы, которые наслаиваются на первичное заболевание (чума свиней, пастереллез и др.) и осложняют его. При этом характерные для сальмонеллеза клинические признаки слабо выражены или отсутствуют, а на вскрытии патолого-анатомические изменения обнаруживаются, главным образом, в органах, где локализуются сальмонеллы (легкие, кишечник, печень).
Краткая историческая справка
Историческая справка, Сальмонеллезы у животных описаны в литературе под названием "паратифы". В 1985 г. Сальмон и Смит выделили из трупов свиней первого представителя обширной группы микробов - Salmonella suipestifer. В дальнейшем аналогичные бактерии были выделены от телят (Гертнер, 1888; Томассен, 1897), мышей (Леффлер, 1890), крыс (Даниш, 1893; В.Н. Исаченко, 1897), из абортированных плодов кобыл (Смит, 1897), цыплят и кур (Реттчер, 1900). В связи с тем что при паратифах выделяли сходные по морфологии и биологическим свойствам палочковидные микробы, международное общество микробиологов в 1934 г. рекомендовало именовать бактерии этой группы сальмонеллами (в честь первооткрывателя Сальмона), а болезни, вызываемые ими - сальмонеллезами. В СССР сальмонеллез телят установили в 1926 г. А.В. Синев и С.К. Беззубец, у овец - П.В. Тавельский (1929), у свиней - А.П. Уранов (1929), Н.А. Михин (1934), Р.А. Цион (1936) и П.Н. Андреев (1937).
Характеристика возбудителяВозбудители - бактерии рода Salmonella отнесены к семейству Enterobacteriaceae. Этот род состоит из 2017 серологических вариантов, объединенных по степени антигенного родства в 52 серогруппы, большинство из которых имеют самостоятельные названия.
У телят сальмонеллез вызывают Salmonella dublin, реже - S. typhimurium и S. enteritidis; у поросят - S. choleraesuis, S. typhisuis, реже - S. typhimurium и Salmonella dublin; у овец - S. abortus ovis и реже - S. typhimurium; у лошадей - S. abortusn equi, реже S. typhimurium; у птиц - S. gallinarum-pullorum, S. typhimurium реже - S. enteritidis, S. Heidelberg, S. anatum, S. Haifa, S. infantis; у пушных зверей - S. typhimurium, S. dublin, S. choleraesuis
Морфология. Независимо от серологического варианта сальмонеллы представляют собой мелкие (1-4x0,5 мкм) палочки с закругленными концами, по Граму окрашиваются отрицательно, спор и капсул не образуют, подвижные (за исключением S. gallinarum-pullorum). Большинство сальмонелл растет на обычных питательных средах с рН 7,2-7,6 при температуре 37 С. В жидкой среде (МПБ) они образуют интенсивное помутнение с обильным серо-белым слизистым осадком; на поверхности среды формируется нежная, разбивающаяся при встряхивании пленка. На поверхности агара (МПА) формируют серовато-голубоватые колонии с ровными краями, различного диаметра (1-5 мм). Ферментативные свойства сальмонелл разнообразны, что зависит от серологического варианта. Общие для них свойства - образование сероводорода, утилизация цитратов; они не разжижают желатин, не ферментируют лактозу.
В основу классификации сальмонелл положена их антигенная структура, установленная Кауфманом и Уайтом (1940), по которой определяют группу и серотип возбудителя. Антигены сальмонелл представлены соматическими О-антигенами, расположенными на поверхности клетки, жгутиковыми Н-антигенами и поверхностными К-антигенами. Для серологической дифференциации многочисленных вариантов сальмонелл используют реакцию агглютинации с монорецепторными 0 - и Н-агглютинирующими сыворотками. Клинически выраженную болезнь у животных вызывают сравнительно немногие серовары сальмонелл.
Культуральные свойства. Являются аэробами или факультативными анаэробами. Оптимальная реакция среды для роста слабощелочная (рН 7,2-7,5), а температура роста 37 °С. Правда, сальмонеллы хорошо растут и при комнатной температуре, и даже не исключается их рост при низких плюсовых температурах (5-8°С). По росту на простом агаре и обычных жидких питательных средах сальмонеллы почти не различимы. На мясо-пептонном агаре гладкие - S-формы этих бактерий образуют круглые, полупрозрачные, выпуклые, иногда со слегка вдавленным центром, и влажные колонии с легким металлическим блеском. Многие серологические типы бактерий рода Salmonella формируют на МПА вокруг колоний четко различимый слизистый вал-феномен валообразования. Феномен валообразования закономерно отсутствует у S. typhimurium, S. abortus ovis, S. pullorum, S. gallinarum. Шероховатые-R-формы имеют вид неровно округленных, шероховатых, тусклых и сухих колоний. На скошенном агаре растут пышно, образуя в конденсационной воде сильную муть, на мясо-пептонном бульоне вызывают равномерное помутнение среды, желатины не разжижают, индола не образуют, молоко не ферментируют.
Общность морфологии и ряда культуральных свойств бактерий рода Salmonеlla не позволяет типизировать их по указанным признакам. Для этого кроме морфологии и культуральных свойств изучают ферментативные свойства и антигенную структуру, в отдельных случаях ставят биологическую пробу на лабораторных животных.
Ферментативные свойства. Ферментативные свойства бактерий обусловлены набором ферментов, отражают определенные условия питания и обмена веществ, свойственные данному виду микроорганизмов в тех или иных условиях внешней среды. Бактерии рода Salmonella характеризуются следующими ферментативными свойствами: не разжижают желатина, не разлагают адонита и не ферментируют сахарозу; подавляющее большинство не расщепляет салицина и не разлагает лактозу, не образует индола. не расщепляет мочевину, не дает реакции Фогес-Проскауера (реакция_на ацетилметилкарбинол); ферментирует (за небольшим исключением мальтозу, маннит, сорбит, расщепляет глюкозу с образованием газа (S. typhi, S.pullorum обычно не образуют газа); дает положительную реакцию с метиловым-красным; утилизирует аммоний и редуцирует нитраты; большинство из них продуцирует сероводород.
Для изучения ферментативных свойств бактерий рода Salmonella обычно используют короткий цветной (пестрый) ряд, состоящий из сред с глюкозой, маннитом, арабинозой, дульцитом, рамнозой (среда Биттера): глицеринофуксиновый бульон (бульон Штерна). Помимо указанных сред для дифференциации серологических типов сальмонелл используют также среды с мальтозой, инозитом, ксилозой; лакмусовое молоко (изменение лакмусового молока при росте сальмонелл позволяет их дифференцировать по способности образовывать кислоту или щелочь). Вместо лакмусового можно использовать обезжиренное молоко с индикатором бромтимоловым синим (1 мл 0,4% -ного раствора в 100 мл молока). Известное значение для дифференциации сальмонелл имеет образование сероводорода культурой. Протеолитические свойства исследуют путем посева изучаемой культуры сальмонелл на МПЖ и молоко.
Ввиду сходства бактерий рода Salmonella с другими микроорганизмами семейства Enterobacteriaceae возникает необходимость их дифференциации. В настоящее время в бактериологической практике широко используют - плотные дифференциально-диагностические питательные среды с лактозой (среды Плоскирева, Эндо, Левина). По способности бактерий ферментировать лактозу сальмонеллы отличают от часто сопутствующей Е. соli, поэтому при исследовании материала на сальмонеллы высев на одну из дифференциально-диагностических сред. На этих средах Е. соli, ферментирующая лактозу с образованием кислоты и изменением цвета индикатора, образует колонии, отличающиеся по цвету от колоний сальмонелл, не ферментирующих лактозу. На среде Эндо бактерии Е. соli дают колонии красного цвета, часто с металлическим блеском, сальмонеллы - бесцветные или бледно-розовые (окрашенные - в цвет среды); на среде Плоскирева Е. соli - колонии оранжево-красного цвета, сальмонеллы - прозрачные или нежно-розовые; на среде Левина Е.соli формируют колонии черного цвета, окруженные ободком, сальмонеллы - прозрачные, нежно-розовые или розовато-фиолетовые. Для дифференциации сальмонелл и культурально сходных штаммов, а также бактерий рода Proteus и бактерий группы кишечных палочек применяют среды с мочевиной (среда Прейса, Ресселя. Олькеницкого), SS-агар (Salmonella-Shigella - агар) и др. Цвет этих сред обусловлен неодинаковой интенсивностью расщепления микроорганизмами азотистых веществ с образованием щелочных продуктов. Бактерии группы кишечных палочек и Proteus (за исключением 0-формы), как правило, на SS-агаре не дают роста, а сальмонеллы растут в виде нежных, бесцветных колоний. Плотные дифференциально-диагностические среды служат лишь для определения принадлежности бактерий к роду Salmonella и отделения их от сопутствующей микрофлоры.
Для наиболее эффективного выделения сальмонелл из патологического материала, содержащего большое количество сопутствующей микрофлоры, препятствующей их росту, используют специальные среды обогащения (Мюллера, Кауфмана и др.). Тетратионовый натрий добавляемый в среду Мюллера, подавляет рост бактерий группы кишечных палочек, но не препятствует развитию сальмонелл. Среда Кауфмана представляёт собой модифицированную среду Мюллера, к которой добавлены раствор бриллиантовой зелени и натуральная бычья желчь. Эти компоненты задерживают рост бактерий группы кишечных палочек и особенно протеуса, но способствуют росту сальмонелл.
Ферментативные свойства сальмонелл не всегда стабильны и могут изменяться в зависимости от условий внешней среды, поэтому правильное типизирование сальмонелл возможно лишь в результате изучения комплекса морфологических, культуральных, ферментативных свойств и антигенной структуры.
Антигенная структура. Она детально изучена Кауфманом, Уайтом, положена в основу современной серологической классификации бактерий рода Salmonella.
У бактерий рода Salmonella различают два основных антигенных комплекса: О - и Н-антигены. Это структурные элементы бактериальной клетки.
Соматические О-антигены термоустойчивы и представляют собой липополисахариднополипептидные комплексы. Жгутиковые Н-антигены термолабильны, имеют белковую природу. Кроме того, у бактерий рода Salmonella обнаружен ряд других антигенов - поверхностных и капсульных. Между капсульными и поверхностными антигенами не существует резкого разграничения, переход осуществляется постепенно, поэтому оба антигена, и капсульный, и поверхностный, объединяются под общим названием К-антиген.
Название К происходит от немецкого слова "kapsel". В группе Salmonella доказано наличие трех К-антигенов: антиген 5, Vi-антиген и М-антиген.
Схема серологической классификации сальмонелл разработана Кауфманом и Уайтом. Согласно предложенной схеме бактерии из рода Salmonella были разбиты на пять больших групп по общности соматического О-антигена: A, В, С, D, E. Оказалось, что O-антигены "неоднородны" и состоят из двух и более рецепторов (фракций), которые были обозначены в схеме римскими цифрами (I, II, III и т.д.). В свою очередь H-антигены, специфические и неспецифические оказались также неоднородными. Рецепторы специфических H-антигенов были обозначены малыми буквами латинского алфавита, а рецепторы неспецифических H-антигенов - арабскими цифрами и частично буквами.
Дальнейшее изучение антигенной структуры бактерий из рода Salmonella, выделенных от людей и животных, обнаруживало все большую сложность этой структуры, сопровождаясь все время открытием новых О - и Н-антигенов, а следовательно, и новых типов. В серологические схемы Кауфмана - Уайта в 1939 г. на II Международном конгрессе микробиологов было введено разделение Н-антигена на фазы I и II с упразднением деления на специфическую и неспецифическую фазы. В обозначении О-антигенов сальмонелл римские цифры были заменены арабскими.
В настоящее время для обозначения серологических групп в схеме исчерпаны все буквы латинского алфавита и последующие группы (51 и дальше) обозначены цифрами их соматических антигенов. Число систематизированных сальмонелл превысило в настоящее время 1600.
Для полного типизирования сальмонелл по антигенной структуре достаточно иметь ограниченный набор монорецепторных О - и Н-сывороток, позволяющих идентифицировать типы сальмонелл групп А, В, С,D, E, которые чаще всего выделяются от людей и животных.
Помимо вышеописанных методов типизирования сальмонелл в последнее десятилетие нашел применение метод фаготипизирования.
В связи с тем что микроорганизмы из рода Salmonella, вызывающие пищевые токсикоинфекции, широко распространены в объектах внешней среды, особенно в пищевых продуктах, важное значение имеют сведения о влиянии на их жизнедеятельность различных физических факторов и химических веществ.
Сальмонеллы обладают свойством продуцировать эндотоксины. Последние термостабильны, представляют собой глюцидо-липоидо-полипептидные комплексы, тождественные с соматическим антигеном бактерий. Многочисленными опытами установлено, что при парентеральном введении они обладают высокой токсичностью. Так, доза в 0,3 мл профильтрованной 7-дневной жидкой культуры при подкожном введении вызывает быструю гибель мышей. В то же время 10-30-кратные дозы, большие предыдущих, при энтеральном введении не вызывали заболевания животных. То же самое было подтверждено в опытах на обезьянах. Наконец, люди добровольно в порядке самоэксперимента выпивали до приема пищи от 20 до 350 мл токсинов сальмонелл (фильтрат убитой культуры), и заболевание у них не наступало. На основании этих опытов был сделан вывод о том, что энтерально действующих токсинов у сальмонелл нет, а пищевые заболевания у людей вызывают только живые бактерии.
Вместе с большой общностью морфологических и культуральных характеристик, а также токсинообразованием бактерии рода сальмонелла отличаются друг от друга по биохимическим и антигенным (серологическим) свойствам. Эти различия и положены в основу научно-разработанных методов типизации.
Устойчивость - в почве, навозе сальмонеллы сохраняются 9-10 мес, в питьевой воде 10-120 дней, в комнатной пыли - 8-18 мес. Переносят замораживание до 4-5 месяцев, при нагревании до 70-75 °С погибают за 15-20 мин; 20 %-ная взвесь свежегашеной извести, содержащая 2% активного хлора, надежно обеззараживают помещения от сальмонелл при экспозиции не менее 1 ч.
Засол и копчение не обеспечивают обеззараживание мяса; проваркой мяса в кусках весом 1 кг в течение 3 ч достигается надежное обеззараживание.
Клинические признаки заболеванияСальмонеллез у молодняка протекает остро, подостро, хронически и атипично (у телят). Инкубационный период колеблется от 1-3 до 7 сут, а его продолжительность зависит от резистентности организма, вирулентности и дозы возбудителя, а также способа заражения и условий, в которых находится восприимчивое животное.
При остром течении у телят, поросят, ягнят и жеребят наблюдается вялость, температура тела повышается на 1-2 °С. Заболевшие больше лежат, дыхание учащенное. Аппетит изменчив, появляется жажда. В день повышения температуры тела, как правило, отмечают диарею (понос). Фекалии становятся жидкими, серо-желтоватого цвета с примесью слизи, пузырьков газа, нередко - крови. В дальнейшем диарея усиливается, и жидкие массы вытекают из ануса непроизвольно. При тяжелом течении поражаются почки: моча становится мутной, иногда с примесью эритроцитов, мочеиспускание частое, болезненное. Наблюдаются артриты: чаще поражаются запястные и заплюсневые суставы.
У поросят в дополнение к указанным признакам развивается конъюнктивит с выделением экссудата, который, засыхая в виде желто-грязных корочек, склеивает веки. На коже области живота, пахов, кончиков ушных раковин появляются очага от темно-синего до фиолетового цвета. У жеребят, заразившихся в период внутри-утробного развития, клинические признаки острого сальмонеллеза проявляются сразу после рождения.
Подострое течение характеризуется менее выраженными симптомами, с появлением признаков пневмонии (истечения из носовых ходов, кашель, хрипы в легких, лихорадка перемежающегося типа), При хроническом сальмонеллезе, который чаще развивается после острой или подострой стадии, наряду с упорными диареями начинают преобладать признаки воспаления легких. Больные-хроники резко отстают в росте, упитанность у них снижается; поражаются запястные, коленные, заплюсневые суставы. У поросят, кроме того, кожа утрачивает эластичность, на ней появляется струпьевидная экзема, кожа ушных раковин темно-фиолетовая с очагами некроза.
Иногда у телят старших возрастных групп (2-4 мес) наблюдают легкое переболевание сальмонеллезом (атипичная, или абортивная, форма). У них уменьшается аппетит, незначительно повышается температура тела. У отдельных появляется понос, и через 3-6 дней животные выздоравливают.
У пушных зверей, заразившихся сальмонеллами, повышается температура тела, отмечают понос и нередко рвоту. При остром течении больные погибают на 2-3-й день, при подостром - на - 14-й день. У самок, заболевших в период гона или беременности, наблюдают бесплодие (14-20%), аборты (до 15%) и большой (до 20%) молодняка в первые 10 дней после щенения.
Инкубационный период и патогенез
Инкубационный период при сальмонеллезах молодняка различный Он зависит от способа заражения, резистентности организма молодняка, вирулентности возбудителя дозы микробных тел и факторов внешней среды
У телят, по данным Н.А. Михина (1936) пои искусственном их инфицировании скрытый период колеблется в пределах 1,5-2 сут и очень редко длится 5 сут. По нашим наблюдениям, он зависит также от пути введения культуры: при подкожном заражении телят в дозе 10 млрд. микробных тел уже через 4-6 ч проявлялись некоторые клинические симптомы заболевания. Иногда при искусственном введении сильновирулентных штаммов внутрь с молоком инкубационный период сокращался до 12 ч. При естественном контактном заражении инкубационный период колеблется в пределах 5-24 сут (Н.А. Михин, 1936).
Экспериментально Н.А. Михин установил, что ежедневные, систематические приемы хотя бы и небольшого количества инфекционного начала приводят к накоплению возбудителя в организме, к перегрузке его патогенными микробами, что вызывает болезнь со смертельным исходом.
В условиях хозяйства А.М. Ахмедов наблюдал инкубационный период длительностью от 4 до 20 дней. Такое различие в днях зависит, как нам кажется, от степени инфицированности сальмонеллами среды, окружающей телят. У телят, выращиваемых на подсосе, инкубационный период болезни был длительным (7 - 20 дней), а в условиях, когда больных телят не изолировали и молоко выпаивали часто из одной и той же посуды, инкубационный период был коротким (4-8 дней). Последнее обусловлено тем, что при ручной выпойке возможность массированного заражения более вероятна, чем при подсосном содержании животных.
У поросят инкубационный период сальмонеллеза как указывает Р.А. Цион (1963), в среднем равен 4-5, а при стационарности болезни-8-12 дням. По Н.А. Михину (1940), он колеблется от 3 до 30 дней. А.Г. Бахтин, связывая продолжительность инкубационного периода с возрастом поросят, сообщает, что у поросят сосунов он бывает в пределах 1-10 дней, а у поросят-отъемышей-до 20 дней. При экспериментальном заражении 11 поросят в дозе 5-10 млрд. S. cholerae-suis внутрибрюшинно все они заболели через 10-24 часа, из них в течение 8 дней пало 10 поросят.
У ягнят и жеребят инкубационный период сальмонеллеза короткий. В опятах А.В. Селиванова (1959) при искусственном заражении ягнят перорально он составлял 2-5 дней и лишь в одном случае-16 дней.
По нашим наблюдения в естественных условиях инкубационный период равнялся 1 - 10 дням. При экспериментальном заражении ягнят различного возраста в дозе от 1 до 3 млрд. микробных тел на 1 кг массы животных инкубационный период у ягнят 4 - 6-дневного возраста был равен 12 - 24 ч, 9 - 12-дневного - 24 - 28ч 20 - 28-дневного - 24 - 72 ч.
По сообщению Е.С. Орлова (1963), ягнята в большинстве случаев заболевали в первые три дня после рождения. Это наблюдение, как и некоторых других авторов, свидетельствует о внутриутробном заражении ягнят сальмонеллезом. В таких случаях очень трудно установить продолжительность инкубационного периода.
У жеребят инкубационный период часто связан с сальмонеллезным абортам кобыл, когда молодняк рождается больным и погибает в первые часы или через 1 - 2 дня после рождения (И.В. Поддубский, 1954) При алиментарном заражении жеребят инкубационный период в среднем равен 3-6 дням.
У птиц инкубационный период болезни колеблется в зависимости от возраста птиц и особенностей возбудителей. Гибель утиных и гусиных эмбрионов при сальмонеллезе чаще всего происходит с 7-го по 22й день эмбрионального развития. При сальмонеллезе кур инкубационный период длится от 12 ч до 7 суток. При пуллороз-тифе у цыплят инкубационный период длится 1-5 дней, а у кур 3-7 дней, иногда дольше.
Патогенез.
Возбудитель, попавший в желудочно-кишечный тракт, быстро проникает в лимфатический аппарат кишечной стенки, а оттуда в лимфо - и кровообращение. Однако размножается ли возбудитель, попавший в желудочно-кишечный тракт, мнения расходятся. Одни авторы считают, что обильного размножения бактерий в кишечнике, по-видимому, не происходит так как в первые дни заболевания обнаружить бактерий в кале удается с трудом. В противоположность этому мнению И.Ф. Квеситадзе (1950) пишет, что микробы Гертнера, попадая в организм животного, начинают быстро размножаться в кишечнике и из него распространяются в другие органы - печень, селезенку, почки и пр., вследствие чего заболевание принимает септицемическую форму. Н.А. Михин (1943) также считает, что сальмонеллы некоторое время размножаются в пищеварительном тракте.
Исследованиями на лабораторных животных, зараженных S. typhimurium, А.А. Вальдман и Е.Н. Ростовой (1955) показали, что у большинства мышей в содержимом желудочно-кишечного тракта уже через 24 ч не удается обнаружить жизнеспособных сальмонелл, так как они из тонкого отдела кишечника постепенно перемещаются в толстый отдел.
П.И. Притулин (1963), изучая патогенез сальмонеллеза на мелких животных с помощью люминесцентного анализа, установил, что возбудитель при пероральном заражении внедряется в организм главным образом через тонкий отдел кишечника.
Из приведенных данных видно, что в первое время после заражения часть возбудителей, попавших в желудочно-кишечный тракт, погибает, часть выделяется с калом во внешнюю среду, а часть внедряется в организм. Скорость внедрения в большой степени зависит от количества эндотоксина, попавшего в организм животных вместе с бактериями. Наблюдения Б.Л. Бамма, М.А. Лебедева (1948), И.В. Шура и Я.П. Шлипакова (1953) показали, что токсины сальмонелл действительно ускоряют внедрение бактерий в организм.
И.В. Шур, Я.П. Шлипаков одной группе мышеи скармливали смесь убитых и живых культур бактерий S. dublin, S. typhimurium, S. cholerae-suis, другой - только живые культуры, а третьей - убитые. Мыши первои группы заболели и погибли на 4 - 5-е сут, а бактерии в их органах обнаруживались через 4-12 ч после заражения; мыши второй группы заболели и погибли на 7-8е сутки, бактерии в их органах обнаруживали через 48-72ч; мыши третьей группы не заболели.
Наблюдения Р.А. Циона (1949) показывают, что массивные дозы инфицированного материала, повышенная вирулентность и токсичность ею прорывает защитную блокаду не только у ослабленных, но и у достаточно сильных телят.
Сальмонеллы в незначительном количестве могут проникнуть в организм через слизистую оболочку ротовой полости и верхнего отдела желудочно-кишечного тракта, в основном же они проникают из тонкого отдела кишечника в лимфатические образования, а именно в солитарные фолликулы и пейеровы бляшки, а также в мезентеральные лимфатические узлы. В них возбудитель размножается и вызывает патологические изменения. Пейеровы бляшки и солитарные фолликулы увеличиваются, отчетливо выступают над слизистой, образуют возвышения, напоминающие по своей форме гряды, вытянутые вдоль кишечника (пейеровы бляшки), или полушаровидные образования (солитарные фолликулы. Из лимфатического аппарата кишечного тракта сальмонеллы могут лопасть в кровь (развивается бактериемия). С поступлением в кровь возбудителя заканчивается инкубационный период и проявляются клинические симптомы болезни.
В первое время л кровь поступает незначительное количество бактерий, но в последующем число их возрастает; этим и объясняется тот факт, что гемокультуры легко удается получить на второй, третий и четвертый день болезни. Часть бактерий и крови разрушается, освобождается эндотоксин.
С кровью возбудитель проникает во все органы, но в наибольшем количестве в те из них, которые богаты Ретикулоэндотелиальними элементами. По мнению И. И, Архангельского (1950), одинаковый процент обнаружения сальмонелл в кале (1,1) и печени (1) косвенно подстверждает циркуляцию возбудителя в организме. Н.И. Притулин (1963) в развитии патологического процесса при сальмонеллезе различает несколько фаз: адаптации микроорганизмов; регионарной инфекции; токсемии; гематогенной дессиминации; септицемии. В фазе регионарной инфекции некоторые бактерии палочковидной формы превращаются в кокковидные и деформированные формы, мелкие палочки и начинает проявляться бактериолизис и фагоцитоз. Люминесцентным анализом автор установил, что фазы регионарной инфекции и последующей токсемии сменяются фазой гематогенной дессиминации (бактериемией);
В патогенезе сальмонеллеза большую роль токсины. А.И. Аверихин (1955) в опытах на установил, что при данной болезни патологические изменения в организме в большинстве случаев развиваются под влиянием сальмонеллезного токсина. Поражение центральной нервной системы при этом носит токсикодистрофический характер. Поражение головного мозга приводит к нарушению высшей нервной деятельности следствием чего является изменение обмена веществ с возникновением нервно-дистрофических процессов в различных органах и тканях.
А.А. Вальдману (1955) при изучении периферической соматической и вегетативной нервной систем, включая интрамуральный отдел тонких кишок, не удалось обнаружить изменений в нервных клетках и нервных волокнах. Однако, как указывает автор, морфологические изменения, обнаруживаемые в клетках центральной нервной системы, являются проявлением сальмонеллез-Ной интоксикации.
М.М. Агабабян (1970) указывает, что при заражении овцематок сальмонеллы не сразу проникают в плод. Вначале под действием токсина, выделяемого 5. аЬог-1из-оУ15, нарушается барьерная функция плаценты матери, затем плода и лишь после этого бактерии проникают в плод.
В.М. Субботин (1955) указывает, что при попадании сальмонелл в организм теленка образуются продукты их жизнедеятельности, которые являются сильными раздражителями чувствительных нервных окончаний и действуют по принципу рефлекса. Исследователь установил ослабление секреторной и моторной деятельности пищеварительного тракта телят и уменьшение переваривающей силы ферментов сычуга и поджелудочной железы. Если организм теленка обладает достаточной резистентностью, то под влиянием защитных факторов (фагоциты, антитела и т.д.) часть возбудителей погибает в крови, а часть выделяется различными путями во внешнюю среду. В организме вырабатывается иммунитет. Поэтому при затяжном течении болезни кровь дает положительную реакцию агглютинации с сальмонеллезным антигеном в титре 1: 100 и выше. Если же резистентность организма слабая, то возбудитель размножается во всех органах и тканях, что вызывает гибель животного.
Резистентность организма имеет большое значение при заболевании птиц сальмонеллезом. Резистентность в свою очередь, зависит от условий кормления и содержания, а также от возраста птиц. Например, возникновению пуллороза способствует перегревание цыплят, индюшат, в результате которого у птенцов происходит торможение функции центральной нервной системы угнетением условных рефлексов и снижением функций всех органов. Кислотно-щелочное равновесие крови изменяется в сторону ацидоза, повышается проходимость для бактерий эпителиального покрова слизистой оболочки кишечника, нарушается деятельность органов пищеварения, тормозится выделение желудочного сока, понижается его кислотность. Сальмонеллы, проникшие в организм, попадают в различные внутренние органы, в том числе и в яичники, а из них проникают в яйца. При инкубировании таких яиц развиваются эмбрионы, пораженные сальмонеллами (пуллороз-тиф цыплят, сальмонеллез эмбрионов водоплавающих птиц).
В местах размножения сальмонелл развивается воспаление (слизистая кишечника, желчный пузырь, печень), а эндотоксин обусловливает экссудативные процессы и диапедез эритроцитов с последующим появлением кровоизлияний на слизистых и серозных оболочках, очаговые некрозы клеток печени, селезенки и почек. Возможны поражения легких, суставов, головного мозга, матки и плода.
В некоторых случаях с момента заражения или после затухания септицемии процесс ограничивается воспалением слизистой оболочки и Лимфатических фолликулов кишечника с образованием изъязвлений и некроза (хронический сальмонеллез). Гибель животного наступает от обезвоживания, многочисленных кровоизлияний, интоксикации и сепсиса.
У переболевших животных возбудитель может локализоваться в желчном пузыре, мезентериальных лимфоузлах и печени, откуда они попадает в просвет кишечника и выделяется во внешнюю среду. У многих клинически выздоровевших животных, вследствие постоянного раздражения слизистой оболочки кишечника, часто отмечают расстройство пищеварения.
Микроорганизмы, попадая в организм животных с кормом или другим путем, вызывают в кишечнике воспалительные процессы, которые обусловливают проникновение возбудителя в кровь и лимфу, и разносятся по всему организму, способствуя развитию септицемии. Микробы и их токсины, циркулируя в крови, вызывают глубокие нарушения в паренхиматозных органах вплоть до образования в них некротических процессов. Последние локализуются в печени, селезенке, почках, легких и суставах. Бактериемия и токсикоз сопровождаются глубокими изменениями эндотелия сосудов микроциркуляторного русла, что ведет к кровоизлияниям, отекам и выходу жидкой части крови в периваскулярные пространства. У беременных животных (лошадей и овец) возбудитель проникает в матку, ткани плода и плодовые оболочки. Патологические процессы в матке обусловливают поражение плода, что сопровождается абортом.
Патологоанатомические изменения.
Патоморофологические изменения при сальмонеллезе у разных видов животных могут отличаться.
Сальмонеллез (паратиф) телят.
Клинико-анатомически различают острое и хроническое течение болезни. Острая болезнь наблюдается в период массовых отелов и свойственна главным образом телятам раннего (2 - 4-неделыюго) возраста. Хроническое течение возникает как результат затяжного течения острой болезни. Спорадические случаи болезни регистрируют и у взрослого крупного рогатого скота.
Патоморфологические изменения.
Макрокартина.
При остром течении болезни основные изменения сосредоточены в кишечнике в виде воспаления, протекающего от слабо выраженного серозно-катарального энтерита до геморрагического. Подобные признаки могут встречаться и в сычуге с образованием эрозий и язв на слизистой оболочке. Кишечное содержимое водянистое, с неприятным запахом, содержит слизь и кровь. Пейеровы бляшки и солитарные фолликулы набухшие и выступают над слизистой оболочкой, на разрезе серо-красного цвета. Если процесс затягивается, в этих участках слизистая оболочка некротизируется и образуется фибринозная пленка. Мезентериальные лимфатические узлы увеличены, отечны, иногда с кровоизлияниями. Селезенка в острых случаях увеличена, пульпа обычно темно-красного цвета, фолликулы не заметны. Этот процесс может смениться острым спленитом - проявляется большое количество беловато-серых узелков. Печень со стертым рисунком, набухшая, дряблая, с множеством мутновато-желтых узелков, достигающих величины булавочной головки, но проявляются они не всегда четко. Стенка желчного пузыря утолщенная и набухшая. Под серозными оболочками в ряде органов могут быть точечно-полосчатые кровоизлияния, при остром заболевании их почти всегда находят под эпикардом и капсулой почек.
В затяжных и хронических случаях часто поражаются легкие. Первоначально в них развивается воспаление типа серозно-катаральной пневмонии, в дальнейшем может быть крупозная пневмония с некротическими очагами. Поражаются преимущественно верхушечная и средние доли, реже - диафрагмальные. Воспаленные доли сине-красного цвета, плотные, влажные на разрезе. Из бронхов выделяется гнойно-катаральная масса. Нередко паратифозная пневмония сопровождается фибринозным плевритом и перикардитом. У взрослых животных поражается также тонкий кишечник, беременные могут абортировать. Основные изменения те же, что и у телят, за исключением более сильно выраженных кровоизлияний на серозных покровах.
Микрокартина.
При гистологическом исследовании в селезенке находят диффузную или очаговую крупноклеточную гиперплазию с некробиотическими изменениями в очагах пролиферации. В печени узелки могут быть двух типов. Одни из них - простые неспецифические токсические некрозы печеночных клеток, другие - гранулемы, расположенные интралобулярно. Они известны под названием паратифозных узелков. Простые некрозы - результат сильного токсического действия на печеночные клетки и эндотелий сосудов продуктов обмена бактерий, представляют собой коагуляционно-некротические очажки. Вокруг них реактивная зона отсутствует или можно наблюдать набухание и пролиферацию клеток ретикуло-эндотелия.
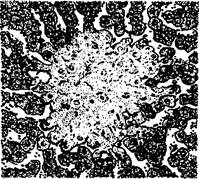
Некротический фокус в печени теленка при сальмонеллезе
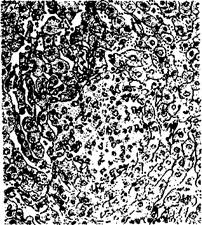
Сальмонеллезная гранулема в печени теленка
Гранулемы (паратифозные узелки) имеют вид очажков размножающихся ретикулоэндотелиальных элементов, расположенных по ходу капилляров. Клетки гранулем со светлыми крупными ядрами обладают способностью к фагоцитозу. Кроме печени и селезенки паратифозные узелки отмечают также в почках, лимфатических узлах, костном мозге. В печени и реже в других органах можно наблюдать образование очаговых эндофлебитов, но они не патогномоничны для сальмонеллеза.
В легких резко выраженный гнойно-катаральный бронхит, одновременно присутствуют признаки катаральной и крупозной пневмонии, чем и объясняется слабая гепатизация пораженных долей. При затяжной пневмонии устанавливают хронический катаральный бронхит и бронхиолит, разрастание соединительной ткани вокруг бронхов, а также в стенках альвеолярных перегородок, что приводит к облитерирующему бронху и