Современные представления о физиологических механизмах сна и сновидений. Организация сна детей и подростков
СОН — состояние относительного покоя, возникающее у животных и человека через определенные промежутки времени, сопровождающееся понижением уровня работы отдельных органов и ряда функции организма. Сон является потребностью всех без исключения животных. Длительное, многодневное лишение сна приводит к ряду тяжелых нарушений в организме и оканчивается смертью. Установлена следующая потребность во сне людей различных возрастов: ребенку до 1 года требуется 16 часов; 3-летнему— 13; в 5 лет— 12, в 7 лет—11,5 в 10—12-летнем возрасте достаточен 10-часовой сон; в 16-летнем — 8 часов, в 17 лет и старше — 8 часов и сутки. Таким образом, взрослый человек проводит в состоянии сна около трети своей жизни. Сопровождающее сон состояние общего покоя имеет важное значение для организма, способствуя восстановлению функции, в первую очередь нервной системы. Нервная система, так же как и другие органы и ткани, во время сна как бы вновь заряжается энергией. Вот почему после глубокого и здорового сна человек испытывает ощущение свежести и бодрости. Исходя из этого, я решила написать курсовую работу, в которой бы рассматривались важные вопросы, находящиеся на стыке нескольких наук, а именно: психологии, физиологии, медицины. Сон - явление очень интересное, важное и сложное. Именно поэтому для его раскрытия нужны знания и конкретные методики вышеперечисленных наук.
Во сне мы проводим около трети жизни. Но сожалеть об этом времени, считать его потраченным впустую, по меньшей мере, несправедливо. Ведь мы спим не только потому, что наше тело нуждается в отдыхе. Пробуждение после крепкого здорового сна можно сравнить с маленьким рождением – обновлением организма. Ведь только благодаря сну мы способны каждый день полноценно работать и активно отдыхать, ясно мыслить и чем-то увлекаться в жизни.
Что же такое сон? Отчего он возникает и насколько велика потребность в нём? И стоит ли придавать такое большое значение сновидениям? Попробуем немного приоткрыть тайну сна и ответить на эти вопросы.
"Кто познает тайну сна, познает тайну мозга". М. Жуве.
1. Научные исследования сна и сновидений
Вопреки извечному человеческому очарованию снами, они не рассматривались в качестве предмета широких научных исследований вплоть до второй половины XX века. Одной из причин этого было то, что научному интересу к процессам сна пришлось ждать возникновения экспериментальной психологии в девятнадцатом веке и развития её в двадцатом. Другой причиной оказался технический фактор: вплоть до недавнего времени инструментарий для исследования снов просто не был разработан. Сложные и чувствительные электронные приборы, используемые в современных исследованиях сна и снов, занимаются измерением, проверкой и записью тончайших нюансов электропотенциала и всех видов биологической активности. До их изобретения у ученых не было возможности отслеживать изменения биоэлектрического потенциала, происходящие в мозгу спящего, сопровождающие (и, может быть, порождающие) события, переживаемые человеком во сне.
В 1875 году Кейтон попытался измерить предполагаемую реакцию мозга на сенсорную стимуляцию. Подвергнув собаку анестезии, он вскрыл ей черепную коробку и обнаружил поверхность полушарии её мозга. Когда Кейтон подсоединил электроды к коре головною мозга собаки, у неё случился шок, и это не был электрошок. Собака была под анестезией, следовательно, возможности получать какую-либо сенсорную информацию у неё не было, и Кейтон не ожидал никаких физиологических изменений в её мозговой активности. По, вопреки ожидаемой стабильности потенциала, в мозгу собаки происходили непрерывные изменения, быстрые колебания напряжения. Произошедшее послужило явным доказательством того, что мозг не является только лишь аппаратом реакций па стимулы: нейтральным его состоянием оказался не полный покой, а активность.(2)
Сведения по проблеме сна были впервые обобщены лишь в 1896 г. М. М. Манасеиной. Первые клинико-морфологические исследования роли поражения отдельных областей мозга в происхождении патологической сонливости принадлежат французскому исследователю Гайе (С. J. A. Gayet, 1875) и австрийскому врачу Маутнеру (L. Mauthner, 1890). Большой вклад в физиологию и патологию состояний бодрствования и сна внес К. Эконома, показавший в 1926 г. на примере летаргического сна при эпидемическом энцефалите значение мезенцефально-гипоталамических структур в поддержании состояний бодрствования и сна. Позднее, в 30—40-х гг. 20 в., в экспериментах на животных было подтверждено важное значение структур мезенцефально-гипоталамического стыка в обеспечении бодрствования и преоптической зоны гипоталамуса — в генезе сна.
Принципиально новым этапом исследований в области проблемы сна явились работы И. П. Павлова и его сотрудников. В соответствии со своим учением о высшей нервной деятельности И. П. Павлов рассматривал сон как разлитое корковое торможение, считая, что внутреннее торможение и сна по физико-химической основе представляют собой один и тот же процесс.
В 1944 г. швейцарский физиолог В. Хесс обнаружил, что электрическое раздражение зрительных бугров вызывало у экспериментальных животных «поведенческий сон», не отличающийся по внешним проявлениям от естественного сна.
Следующий этап развития представлений о механизме сна связан с анализом роли ретикулярной формации ствола в механизмах деятельности мозга. В исследованиях Дж. Моруцци и X. Мегуна (1949) было обнаружено важнейшее значение восходящих активирующих влияний ретикулярной формации ствола мозга и гипоталамуса на вышележащие отделы в поддержании бодрствования. Сон при этом рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включенном» таламокортикальных синхронизирующих процессов. Несколько позднее была показана роль каудальных отделов мозгового ствола в возникновении сна. Эти исследования положили начало развитию представлении об активной природе сна, что затем было подтверждено в опытах на животных, а также на человеке.(3)
Ханс Бергер, немецкий нейропсихиатр, при помощи новых приборов записал электрическую активность человеческого мозга. Результаты оказались не менее сенсационными, чем открытие, сделанное Кейтоном за 50 лет до него. В опытах с человеком Бергер ожидал получить такие же беспорядочные колебания напряжения, как и при проведении опытов с животными: кроликами, кошками, собаками, обезьянами. Но колебания напряжения у представителей человеческой расы оказались неожиданно ритмичными. Бергер назвал записи мозговых волн электроэнцефалограммой (ЭЭГ) и отметил, что, как только субъект был в состоянии лечь, закрыть глаза и расслабиться, колебания его мозговых волн становились регулярными, с периодичностью повторения примерно 10 раз в секунду. Это и был знаменитый "альфа-ритм" (названный так его первооткрывателем), свидетельствующий о состоянии расслабления (равно как и о погружении в медитацию). Бергер обнаружил, что частота (количество пиков в секунду) колеблется между 8-ю и 12-ю, и альфа-ритм исчезает, как только из внешнего мира поступает неожиданный стимул (например, звук щелчка пальцами). Наконец-то у науки появилось окно, открытие которого обещало пролить свет на природу сознания.
Но лишь после повторения этого опыта исследователями из Кембриджского университета, основополагающее открытие Бергера было принято, и тем самым положено начало энцефалографии как науки. Среди исследований связи между состоянием сознания и состоянием мозга (в которых Бергер также был пионером) была и первая электроэнцефалограмма спящего человека.
Исследования изменений ЭЭГ в процессе сна, впервые выявленных Бергером, были продолжены в 30-е годы в Гарвардском университете. На основе записей ЭЭГ бодрствования и сна пяти уровней там пришли к заключению, что сновидения имеют место во время более поверхностного сна. В подобной же серии исследовании в Чикагском университете изучалась разница между изменениями умственной активности у бодрствующего и у спящего субъекта. Был сделан вывод, что в фазе глубокого сна сны снятся очень редко. Эти исследования позволили предположить, что изучение сновидения могло бы стать более объективным и научным, если бы существовали какие-то способы удостовериться, видит данный человек сны, или нет - и если видит, то когда. Но прежде чем ученые реализовали эту возможность, прошло несколько десятилетий.
В конце 40-х было обнаружено, что стимуляция нервной структуры ствола мозга (основания мозга), называемой ретикулярной формацией, ведет к активизации коры больших полушарий. Стимуляция ретикулярной формации у спящей кошки, к примеру, приводила к пробуждению, а разрушение приводило, наоборот, к состоянию перманентной комы. А коль скоро главным источником активизации ретикулярной формации являются сенсорные сигналы, была предложена теория, согласно которой сон может порождать процессы торможения в ретикулярной системе. Так что погружение в сон может зависеть от снижения ретикулярной активности вследствие уменьшения количества поступающих сенсорных сигналов.
Засыпание не могло быть объяснено только этой теорией. Поэтому обнаружение через некоторое время в основании мозга, лобных долях и других его частях активных гипногенных центров, электро - или нейрохимическая стимуляция которых вела к засыпанию, не было неожиданностью.
К концу 40-х годов это было существенным достижением в научном изучении биологии сна. Сон рассматривался как конец континуума бодрствования. В другом конце этого континуума было состояние полного бодрствования, поделенное на промежуточные стадии: от расслабления, через состояние внимания и до состояния полной умственной подвижности, достигающей крайней степени в маниях или в панике. В каком месте этой шкалы вы находитесь, зависит от состояния вашей ретикулярной формации. При таком подходе сон становится банальностью, и степень погружения в него определялась по шкале бодрствования. Сновидения, отмечавшиеся чаще всего во время неглубокого сна, выглядели как занятные отклонения в сторону состояния частичного бодрствования при частичном функционировании аппарата мышления.
С течением времени эти взгляды были вытеснены новыми, возникшими в результате важных событий 50-х годов.
В 1953 году студент Чикагского университета Юджин Азеринский (Е. Aserinsky), работая под руководством Натаниэля Клейтмана (N. Kleitman), изучал картину сна у младенцев и сделал важное наблюдение. Он заметил, что периоды движения глаз и других проявлений активности регулярно перемежаются периодами относительно спокойного сна. Эти повторяющиеся периоды быстрых движений глаз (БДГ) легко наблюдать при помощи электродов, прикрепленных у глаз наблюдаемого. Получившуюся запись назвали электроокулограммой (ЭОГ). Одновременное снятие записей ЭЭГ и ЭОГ показало, что периоды БДГ соответствуют поверхностному сну. Больше того, когда субъекты (в данном случае - взрослые) бывали разбужены после БДГ-периодов, они обычно сообщали об очень ярких снах; после же пробуждения от других фаз сна они рассказывали, что видели сны только в одном случае из пяти (другие фазы называются не-БДГ-сон, или НБДГ).
Наконец-то у науки появился ключ к снам или, по крайней мере, к разрешению таких загадок, как частота и длительность сновидений, а также вопроса о том, бывают ли люди, невидящие снов, или они их просто не помнят.
Среди тех, кто работал в лабораториях Клейтмана, был медик-второкурсник Уильям К. Демент, защитивший по окончании медицинского колледжа докторскую диссертацию по физиологии под руководством Клейтмана. Демент для своей диссертации провел обширную серию экспериментов, направленных на дальнейшее прояснение того, какое отношение имеет БДГ-сон к сновидениям (БДГ-сон - термин, предложенный Дементом). Новые исследования Демента обнаружили множество основных характеристик БДГ-снов. Среди прочего было открытие связи между продолжительностью БДГ-сна перед пробуждением и длиной последующего отчета о сновидении: чем больше времени проведено в БДГ-сне, тем длиннее сновидение. Это дало первое (хотя и косвенное) доказательство существования соответствия между физическим временем и временем сновидения. Демент также предположил, что существует соответствие между движениями глаз и изменением направления взгляда во сне. Предположение, что БДГ являются следствием движений глаз спящего в его сновидении, вызвало в дальнейшем значительные дискуссии.(5)
Я хочу отметить, что следствием основополагающих работ Азеринского, Демента и Клейтмана в последующие 30 лет стали тысячи исследований сна и сновидений.
2. Физиологические механизмы сна и сновидений
Прежде всего, заметим, что сон – это естественное состояние организма, сила его неодолима. Природа сна так до сих пор и не понята окончательно, хотя мы уже рассмотрели немало попыток разобраться в физиологии этого явления.
Сон — естественный физиологический процесс пребывания в состоянии с минимальным уровнем активного сознания и пониженной реакцией на окружающий мир, присущий млекопитающим, птицам, рыбам и некоторым другим животным, в том числе насекомым (например, дрозофилам).
Существуют различные теории, объясняющие причины и механизмы сна.
Гуморальная теория, в качестве причины сна рассматривает вещества, появляющиеся в крови при длительном бодрствовании. Доказательством этой теории служит эксперимент, при котором бодрствующей собаке переливали кровь животного, лишенного сна в течение суток. Животное-реципиент немедленно засыпало. В настоящее время удалось идентифицировать некоторые гипногенные вещества, например пептид, вызывающий дельта-сон. Но гуморальные факторы не могут рассматриваться как абсолютная причина возникновения сна. Об этом свидетельствуют наблюдения за поведением двух пар неразделившихся близнецов. У них разделение нервной системы произошло полностью, а системы кровообращения имели множество анастомозов. Эти близнецы могли спать в разное время: одна девочка, например, могла спать, а другая бодрствовала.
Подкорковая и корковая теории сна. При различных опухолевых или инфекционных поражениях подкорковых, особенно стволовых, образований мозга, у больных отмечаются различные нарушения сна - от бессонницы до длительного летаргического сна, что указывает на наличие подкорковых центров сна. При раздражении задних структур субталамуса и гипоталамуса животные засыпали, а после прекращения раздражения они просыпались, что указывает на наличие в этих структурах центров сна.
В лаборатории И.П.Павлова было установлено, что при длительной выработке тонкого дифференцировочного торможения животные часто засыпали. Поэтому ученый рассматривал сон как следствие процессов внутреннего торможения, как углубленное, разлитое, распространившееся на оба полушария и ближайшую подкорку торможение (корковая теория сна).
Однако ряд фактов не могли объяснить ни корковая, ни подкорковая теории сна. Наблюдения за больными, у которых отсутствовали почти все виды чувствительности, показали, что такие больные впадают в состояние сна как только прерывается поток информации от действующих органов чувств. Например, у одного больного из всех органов чувств был сохранен только один глаз, закрытие которого погружало больного в состояние сна. Многие вопросы организации процессов сна получили объяснение с открытием восходящих активирующих влияний ретикулярной формации ствола мозга на кору больших полушарий. Экспериментально было доказано, что сон возникает во всех случаях устранения восходящих активирующих влияний ретикулярной формации на кору мозга. Были установлены нисходящие влияния коры мозга на подкорковые образования. В бодрствующем состоянии при наличии восходящих активирующих влияний ретикулярной формации на кору мозга нейроны лобной коры тормозят активность нейронов центра сна заднего гипоталамуса. В состоянии сна, когда снижаются восходящие активирующие влияния ретикулярной формации на кору мозга, тормозные влияния лобной коры на гипоталамические центры сна снижаются.
Между лимбико-гипоталамическими и ретикулярными структурами мозга имеются реципрокные отношения. При возбуждении лимбико-гипоталамических структур мозга наблюдается торможение структур ретикулярной формации ствола мозга и наоборот. При бодрствовании за счет потоков афферентации от органов чувств активируются структуры ретикулярной формации, которые оказывают восходящее активирующее влияние на кору больших полушарий. При этом нейроны лобных отделов коры оказывают нисходящие тормозные влияния на центры сна заднего гипоталамуса, что устраняет блокирующие влияния гипо-таламических центров сна на ретикулярную формацию среднего мозга. При уменьшении потока сенсорной информации снижаются восходящие активирующие влияния ретикулярной формации на кору мозга. В результате чего устраняются тормозные влияния лобной коры на нейроны центра сна заднего гипоталамуса, которые начинают еще активнее тормозить ретикулярную формацию ствола мозга. В условиях блокады всех восходящих активирующих влияний подкорковых образований на кору мозга наблюдается медленно-волновая стадия сна.
Гипоталамические центры за счет связей с лимбическими структурами мозга могут оказывать восходящие активирующие влияния на кору мозга при отсутствии влияний ретикулярной формации ствола мозга. Эти механизмы составляют корково-подкорковую теорию сна (П.К.Анохин), которая позволила объяснить все виды сна и его расстройства. Она исходит из того, что состояние сна связано с важнейшим механизмом - снижением восходящих активирующих влияний ретикулярной формации на кору мозга. Сон бескорковых животных и новорожденных детей объясняется слабой выраженностью нисходящих влияний лобной коры на гипоталамические центры сна, которые при этих условиях находятся в активном состоянии и оказывают тормозное действие на нейроны ретикулярной формации ствола мозга.
Сон новорожденного периодически прерывается только возбуждением центра голода, расположенного в латеральных ядрах гипоталамуса, который тормозит активность центра сна. При этом создаются условия для поступления восходящих активирующих влияний ретикулярной формации в кору. Эта теория объясняет многие расстройства сна. Бессонница, например, часто возникает как следствие перевозбуждения коры под влиянием курения, напряженной творческой работы перед сном. При этом усиливаются нисходящие тормозные влияния нейронов лобной коры на гипоталамические центры сна и подавляется механизм их блокирующего действия на ретикулярную формацию ствола мозга. Длительный сон может наблюдаться при раздражении центров заднего гипоталамуса сосудистым или опухолевым патологическим процессом. Возбужденные клетки центра сна непрерывно оказывают блокирующее влияние на нейроны ретикулярной формации ствола мозга.
Иногда во время сна наблюдается так называемое частичное бодрствование, которое объясняется наличием определенных каналов реверберации возбуждений между подкорковыми структурами и корой больших полушарий во время сна на фоне снижения восходящих активирующих влияний ретикулярной формации на кору мозга. Например, кормящая мать может крепко спать и не реагировать на сильные звуки, но она быстро просыпается даже при небольшом шевелении ребенка. В случае патологических изменений в том или ином органе усиленная импульсация от него может определять характер сновидений и быть своего рода предвестником заболевания, субъективные признаки которого еще не воспринимаются в состоянии бодрствования.
Фармакологический сон неадекватен по своим механизмам естественному сну. Снотворные препараты ограничивают активность разных структур мозга - ретикулярной формации, гипоталамической области, коры головного мозга. Это приводит к нарушению естественных механизмов формирования стадий сна, нарушению процесса консолидации памяти, переработки и усвоения информации.(4)
Сон — особое состояние сознания человека и животных, включающее в себя ряд стадий, закономерно повторяющихся в течение ночи. Появление этих стадий обусловлено активностью различных структур мозга.
Изучение торможения показало, что оно не просто препятствует дальнейшей работе нервных клеток. Во время этого внешне пассивного состояния (именно только внешнее, ибо в это самое время внутри клетки совершаются активные процессы обмена веществ) клетки мозга восстанавливают нормальный состав, набирают силы для дальнейшей активной работы. Во сне, когда заторможена подавляющая масса мозга, создаются наиболее благоприятные условия не только для восстановления работоспособности нервных клеток мозга, более всего нуждающихся в такой передышке, но и для отдыха всего организма.
Во время сна у человека периодически чередуются две основные фазы: медленный и быстрый сон, причём в начале сна преобладает длительность медленной фазы, а перед пробуждением — растёт длительность быстрого сна. Полисомнография (система регистрации электроэнцефалограммы, электроокулограммы и электромускулограммы) показывает, что сон у большинства людей состоит из 4—6 волнообразных циклов, длительностью 80 — 100 мин.
Каждый цикл включает фазы «медленного», или ортодоксального, сна (МС), на долю которого приходится 75 % сна, и «быстрого», или парадоксального (БС), составляющего около 25 %. Эти названия обусловлены характерными особенностями ритмики электроэнцефалографии во время сна — медленной активностью в МС и более быстрой — в БС.
При спокойном сне тело спящего неподвижно, глаза закрыты, мышцы расслаблены, дыхание замедлено, контакт с окружающим отсутствует, но во всех частях, органах и системах организма в это время совершаются активные, жизненно важные процессы, способствующие его самообновлению.
В течение 7-8 часового ночного сна мозг проходит циклы глубокого сна, длящиеся в среднем от 30 до 90 минут, за которыми следуют 10-15 минутные эпизоды быстрого сна. К концу ночи, если человека не тревожить, продолжительность медленного сна уменьшается, а количество эпизодов быстрого сна увеличивается.
Когда человек засыпает, обычные для него в состоянии бодрствования мозговые волны плавно сменяются медленными волнами. По мере углубления сна постепенно замедляется частота мозговых волн и увеличивается их амплитуда. Достижение глубокого сна занимает 30-45 минут, после чего процесс оборачивается вспять, занимая 30-45 минут для возвращения в стадию лёгкого сна. В течение этой фазы мышцы сохраняют тонус, а скорость сердцебиения и дыхания замедляется незначительно. Во время быстрого сна глаза под закрытыми веками быстро перемещаются. Мышцы полностью теряют тонус, но мышцы конечностей и лица дёргаются в такт движениям глаз. Дыхание, сердцебиение и кровяное давление подвержены нерегулярным изменениям. Во время быстрого сна у мужчин возникает эрекция, даже у тех, кто не может достичь эрекции в нормальных условиях вследствие нервного расстройства. Если человека разбудить в этой стадии, он говорит о том, что видел сон.(6)
Медленный сон (медленноволновой сон, ортодоксальный сон, ФМС).
Медленный сон разделяется на 4 стадии, отличающиеся биоэлектрическими (электроэнцефалографическими) характеристиками и порогами пробуждения, являющимися объективными показателями глубины сна.
· Первая стадия (дремота). Это переходное состояние между пассивным бодрствованием и легким сном. Характеризуется отсутствием на ЭЭГ альфа-ритма, являющегося характернейшим признаком бодрствования здорового человека, с десинхронизацией (снижением амплитуды) и появлением низкоамплитудной медленной активности с частотой 3 — 7 в 1 сек. (тета- и дельта-ритмы). Могут регистрироваться ритмы и с более высокой частотой. На электроокулограмме возникают изменения биопотенциала, отражающие медленные движения глаз. Поведение: дремота с полусонными мечтаниями и сноподобными галлюцинациями - гипнагогическими (вводящими в сон, по-гречески) образами. В этой стадии могут интуитивно появляться идеи, способствующие успешному решению той или иной проблемы.
· Вторая стадия (сон средней глубины) характеризуется ритмом «сонных веретён» — сигма-ритмов, которые представляют собой учащённые альфа-ритмы (12—14—20 Гц). Их частота 13 — 16 в 1 сек., т. е. отдельные колебания биопотенциалов группируются в пачки, напоминающие форму веретена (рис., в). В этой же стадии из фоновой активности четко выделяются 2 — 3-фазных высокоамплитудных потенциала, носящих название К-комплексов, нередко связанных с «сонными веретенами». К-комплексы регистрируются затем во всех стадиях медленного сна. Амплитуда фоновой ритмики ЭЭГ при этом растет, а частота ее уменьшается по сравнению с первой стадией. С появлением «сонных веретён» происходит отключение сознания; в паузы между веретёнами человека легко разбудить. Повышаются пороги восприятия. Самый чувствительный анализатор — слуховой (мать просыпается на крик ребёнка, каждый человек просыпается на называние своего имени).
· Третья стадия. Для третьей стадии характерно появление на ЭЭГ медленной ритмики в дельта-диапазоне (т. е. частотой до 2 в 1 сек. и амплитудой 50—75 мкв и выше). При этом продолжают достаточно часто возникать «сонные веретена». Стадия глубокого, лишенного сновидений сна.
· Четвёртая стадия. Четвертая стадия (поведенчески наиболее глубокий сон) характеризуется доминированием на ЭЭГ высокоамплитудного медленного дельта-ритма (2 Гц). Третью и четвёртую стадии часто объединяют под названием дельта-сна. В это время человека разбудить очень сложно; возникают 80 % сновидений, и именно на этой стадии возможны приступы лунатизма и ночные кошмары, однако человек почти ничего из этого не помнит. Первые четыре медленноволновые стадии сна в норме занимают 75—80 % всего периода сна.(3)
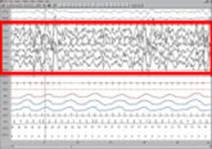
4-я стадия медленного сна, глубокий сон. ЭЭГ выделена красной рамкой.
После погружения в дельта-сон и пребывании в нем тридцать-сорок минут вы возвращаетесь обратно во вторую стадию. Примерно через тридцать-сорок минут после засыпания, вы первый раз за ночь входите в фазу быстрого сна. После 5-10 минут быстрого сна вы, возможно, проснетесь, запомнив сон, и провалитесь обратно во вторую стадию и, возможно, в дельта-сон, наступающие примерно каждые 90 минут перед очередным периодом быстрого сна, и так далее.(1)
ФБС отличается низкоамплитудной ритмикой ЭЭГ, а по частотному диапазону наличием как медленных, так и более высокочастотных ритмов (альфа- и бета-ритмов). Характерными признаками этой фазы сна являются и так называемые пилообразные разряды с частотой 4 в 1 сек., быстрые движения глаз на электроокулограмме, в связи с чем эту фазу часто называют сном с быстрыми движениями глаз, а также резкое снижение амплитуды электромонограммы или полное падение тонуса мышц диафрагмы рта и шейных мышц. Это напоминает состояние бодрствования. Вместе с тем (и это парадоксально!) в эту стадию человек находится в полной неподвижности, вследствие резкого падения мышечного тонуса. Однако глазные яблоки очень часто и периодически совершают быстрые движения под сомкнутыми веками. Существует отчетливая связь между быстрым движением глаз и сновидениями. Если в это время разбудить спящего, то в 90 % случаев можно услышать рассказ о ярком сновидении.
Фаза быстрого сна от цикла к циклу удлиняется, а глубина сна снижается. Быстрый сон прервать труднее, чем медленный, хотя именно быстрый сон ближе к порогу бодрствования.
Прерывание быстрого сна вызывает более тяжёлые нарушения психики по сравнению с нарушениями медленного сна. Часть прерванного быстрого сна должна восполняться в следующих циклах.
Предполагают, что быстрый сон обеспечивает функции психологической защиты, переработку информации, ее обмен между сознанием и подсознанием.
Слепым от рождения во время быстрого сна снятся звуки и ощущения, быстрого движения глаз у них нет.(3)
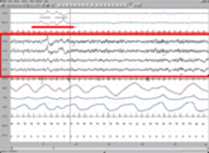
Быстрый сон. ЭЭГ выделена красной рамкой. Движения глаз подчёркнуты красным.
Переход от медленного сна к быстрому весьма отчетлив. Во время быстрого сна, обычно именуемого фазой быстрых движений глаз, или БДГ-сном, ваши зрачки под сомкнутыми веками совершают быстрые движения, почти как в бодрствующем состоянии. Дыхание становится частым и прерывистым, мозг расходует столько же энергии, как при бодрствовании, и вам снятся яркие сны. Но несмотря на мозговую активность, ваше тело остается почти полностью неподвижным (за исключением небольших подергиваний), поскольку во время БДГ-сна оно временно парализуется, чтобы вы не смогли перенести свои действия за пределы сна.(5)
Сон человека и животных циклически организован. У человека длительность одного цикла сна составляет в среднем 1,5—2 часа. За ночь наблюдается 3—5 циклов. Каждый из циклов состоит из отдельных стадий ФМС и ФБС. Первое появление ФБС происходит через 1—1,5 часа после засыпания вслед за стадиями ФМС. Дельта-сон характерен для первых двух циклов сна, длительность же ФБС максимальна на протяжении III и IV циклов (обычно это ранние утренние часы). В среднем у человека в молодом и среднем возрасте ФМС составляет 75—80% длительности всего сна, при этом первая стадия — около 10%, вторая стадия — 45—50% и дельта-сон — около 20%. ФБС занимает соответственно 20—25% длительности сна. Эти значения существенно отличаются от аналогичных показателей у новорожденных детей, а также в пожилом и старческом возрасте.(4)
Во время разных фаз и стадий сна происходят существенные перестройки в деятельности мозга в целом, его отдельных функциональных систем и висцеральных, эндокринных функций. Исследование деятельности отдельных нейронов во время ФМС показало, что средняя частота импульсации в большинстве структур мозга уменьшается, хотя в некоторых из них, активно обеспечивающих наступление и протекание сна, она увеличивается по сравнению с состоянием бодрствования. Изменяется не только частота, но и сам характер нейронной активности. В зрительных буграх и коре мозга взамен постоянной импульсации в состоянии бодрствования во время ФМС импульсы генерируются сериями - последующим периодом молчания (импульсация типа пачка—пауза). Во время ФБС активность нейронов большинства отделов мозга усиливается, достигая уровня бодрствования или даже превосходя ее. Ритм типа пачка—пауза вновь сменяется более постоянной нейронной активностью. Изменяется и возбудимость нейронов; в ФМС по сравнению с бодрствованием она уменьшается и еще более снижается в ФБС.
Для ФБС характерно возникновение своеобразного электрофизиологического феномена — спайков, возникающих в варолиевом мосту и распространяющихся затем в коленчатые тела и зрительную кору головного мозга, где представлен зрительный анализатор. По времени они связаны с появлением быстрых движений глаз.
В период сна происходит существенное уменьшение активности двигательной системы, что обусловлено активным торможением, исходящим из ствола мозга. Выраженность спиральных и бульбарных моносимпатических и полисинаптических двигательных рефлексов снижается в ФМС и еще более в ФБС. Эти изменения, обусловленные пресинаптическим и постсинаптическим торможением деятельности гамма- и альфа-мотонейронов, приводят к снижению мышечного тонуса в ФМС и его резкому подавлению в мышцах головы и шеи на протяжении ФБС. Несмотря на общее снижение двигательной активности, во время сна отмечается наличие различных движений — от мелких (в виде подергивания мышц лица, туловища и конечностей, возникающих при засыпании и учащающихся в период ФБС) до более массивных (в форме перемены позы в постели), наблюдающихся во всех стадиях сна и часто предваряющих смену стадий.
Для сна характерно существенное изменение состояния вегетативно-висцеральной сферы организма. Мозговой кровоток в ФМС существенно не изменяется по сравнению с бодрствованием, а лишь усиливается в некоторых структурах. В ФБС он значительно увеличивается, превосходя показатели спокойного бодрствования, и одновременно повышается и температура мозга. Эти данные, как и характеристика нейронной активности, указывают на высокую функциональную активность мозга во время сна.
Принятая раньше формула «сон - царство вагуса» оказалась верной лишь отчасти. При засыпании и во время первых стадий ФМС действительно снижается артериальное давление, уменьшается частота сердечных сокращений и урежается дыхание. При этом могут быть и физиологические аритмии пульса и дыхания. В более глубоких стадиях ФМС частота сердечных сокращений и дыхания несколько повышается, что возможно носит компенсаторный характер и необходимо для поддержания оптимального уровня системного кровотока и легочной вентиляции в связи со снижением артериального давления и уменьшением глубины дыхания. В ФБС показатели деятельности сердечнососудистой и дыхательной систем резко усиливаются. При этом наряду с повышением артериального давления, частоты сердечных сокращений и дыхания отмечается их большая динамичность, проявляющаяся заметными аритмиями пульса, дыхания.
Фазовые изменения вегетативных функций часто связаны во времени с появлением быстрых движений глаз в ФБС и К-комплексов или активных движений тела в ФМС.
Параллельно с характерными изменениями на ЭЭГ сон сопровождается своеобразной динамикой другого вегетативного показателя — кожно-гальванической реакции, которая, уменьшаясь при засыпании, вновь усиливается в глубоких стадиях ФМС и опять заметно тормозится в ФБС. Во время сна также наблюдается определенная динамика величин электрокожного сопротивления, потоотделения, кожной температуры. Характерным вегетативным феноменом ФБС является эрекция полового члена у мужчин и клитора у женщин, наблюдающаяся в этой фазе сна, как у младенцев, так и у пожилых людей.(1)
В период сна происходят существенные изменения в деятельности эндокринной системы. Следует учитывать, что, например, уменьшение выделения АКТГ и кортизола в вечерние часы и в начале ночи, а также изменения в некоторых других показателях деятельности эндокринной системы связаны непосредственно с циркадианным ритмом, а не с механизмами сна.
Динамика секреции других гормонов (соматотропный гормон и пролактин) имеют большую связь со сном. Пик суточной секреции соматотропного гормона приходится на период дельта-стадии ФМС в I цикле сна. Приблизительно в этот же период наблюдается и один из пиков секреции пролактина, последний из которых приходится на ранние утренние часы.
Психическая деятельность в отдельных стадиях и фазах сна также имеет свою специфику. Для стадии дремоты характерны своеобразные зрительные образы (гипнотические грезы). При пробуждении людей из более глубоких стадий ФМС нередко можно получить отчеты о мыслеподобной психической деятельности, иногда о расплывчатых зрительных образах, не обладающих той яркостью, эмоциональностью, которые характерны для типичных сновидений, возникающих в ФБС.
Несмотря на то, что предположение о наличии так называемых центров сна не подтвердилось, известен целый ряд образований головного мозга, активная деятельность которых обеспечивает возникновение и протекание сна как физиологического процесса. В области продолговатого мозга и варолиевого моста расположены группы клеток, деятельность которых вызывает поведенческий сон и возникновение соответствующей биоэлектрической активности на ЭЭГ. Активность этой зоны тормозит деятельность ретикулярной формации среднего мозга, обеспечивающей состояние бодрствования. Другим важным звеном синхронизирующей (сомногенной) системы мозга является преоптическая область гипоталамуса. Она функционирует синергично с бульбарной ингибиторной зоной и также тормозит деят