Возможное адаптивное значение открытого таза птиц и новая гипотеза происхождения полета
С.А. Лопарев
г. Киев
Пути происхождения класса птиц, наиболее существенных их морфологических приспособлений (перьевой покров, характерная забота о потомстве, полет) — один из самых сложных и запутанных вопросов филогении. Он осложняется еще и крайней немногочисленностью палеонтологических остатков. Большинством исследователей, в основном палеонтологов и эволюционистов, для решения вопроса чаще всего разбираются морфологические особенности, в первую очередь скелета, как современных птиц, так и возможных родственных вымерших форм (Дементьев, 1965; Рауп, Стенли, 1974; Коуэн, 1982; Курзанов, 1987; Грант, 1991; Кэрролл, 1993). Реже, в основном орнитологами, анализируются адаптивные возможности, морфология и экология современных форм. Анализируются количественные соотношения видов птиц различной экологии внутри отрядов, обычно принимаемых как более примитивные, и между отрядами, а также сравнение немногих известных вымерших форм с современными (Познанин, 1957; Дементьев, 1965; Татаринов, 1980). До последнего времени недооценивалось существенное различие между классами птиц и рептилий по уровню метаболизма, степени развития головного мозга и высшей нервной деятельности, весьма существенно влияющим на экологию и поведение форм. Кроме этого, значительную неопределенность в вопрос вносят существенные различия во взглядах на энергетику, экологию и эволюцию морфологических особенностей мезозойских рептилий (Рауп, Стенли, 1974; Коуэн, 1982; Кэрролл, 1993).
Происхождение характерного строения таза
Гипотезы-обоснования
Для всех современных птиц характерно наличие так называемого “открытого” таза и образование сложного синсакрума. Наиболее характерные черты этой структуры — срастание удлиненных подвздошных костей с многими позвонками, разворот их параллельно позвоночнику в почти горизонтальное положение и образование “крыши” тазовой полости. Седалищные кости из обычного рептильного (сохраняюшегося и у млекопитающих) положения отодвигаются в стороны и назад и в подавляющем большинстве прирастают к подвздошным. Симфиз между их направленными назад и вниз отростками отсутствует. Лобковые кости из обычного для рептилий и млекопитающих передне-нижнего положения отодвигаются резко назад и слегка в стороны, занимая положение параллельно седалищным. У значительной части птиц они в большей или меньшей степени срастаются с седалищными. Симфиз между ними тоже отсутствует, исключение — африканский страус (Struthio camelus). Однако, будучи более длинными, чем седалищные, лобковые кости сохраняют свободные концы (у большинства видов с небольшими расширениями), которые могут в зависимости от физиологического состояния особи несколько менять свою ориентацию. В подавляющем большинстве руководств (Шимкевич, 1922; Шмальгаузен, 1938; Наумов, Карташев, 1979; Кэрролл, 1993) такое строение таза объясняется необходимостью откладки крупных яиц и прохождением их через выводные каналы в твердой скорлупе. В некоторых из работ (Шмальгаузен, 1938; Наумов, Карташев, 1979; Коуэн, 1982; Кэрролл, 1993) признается возможным удобство подобной структуры при двуногом передвижении, а в последнее время предполагается связь с интенсификацией дыхания — неограниченная подвижность брюшной стенки (Наумов, Карташев, 1979).
Признавая значение деталей строения птичьего таза для всех этих процессов, мы, тем не менее, склонны объяснять происхождение и формирование его иной причиной, причем признаем ее ведущей. Только впоследствии сформированный птичий таз способствовал выполнению всех вышеперечисленных функций. Причина эта — откладка яиц с чрезвычайно большим количеством желтка, особенно у более примитивных выводковых птиц и втягивание значительных количеств остаточного желтка перед вылуплением через пупочный канал в брюшную (тазовую) полость, где этот желток в первые несколько суток жизни занимает до 2/3 объема, составляя до 30–38 % веса суточного птенца, например, у куриных. Морфоэкологические связи этого явления, его влияние на энергетику ранних стадий и особенности роста мы рассмотрим ниже, а перед этим попробуем проанализировать правомочность альтернативных объяснений.
Необходимость иметь развернутые назад седалищные и лобковые кости для прохождения по родовым каналам крупных яиц в твердой скорлупе не выдерживает критики, поскольку относительный вес (объем) яиц у большинства птиц составляет от 5 до 15 % веса тела, что примерно сответствует относительному весу новорожденных млекопитающих, у части из которых (человек!) имеются значительных объемов голова, или длинные и неудобные при рождении конечности (копытные). Оставляя в стороне китообразных, вес новорожденных у которых может быть до трети веса тела матери (таз китообразных редуцирован), и подковоносов из рукокрылых, у которых вес новорожденных может доходить до 30 % и более веса самки (у подковоносов редуцированы лобковая и седалищная кости) (Бобринский и др., 1965; Мосияш, 1985), остановимся на остальных рукокрылых, имеющих нормальный таз закрытого типа (тазовое кольцо) с лобковым и седалищным симфизами, и тем не менее рождающих детенышей, составляющих в среднем около 20 % веса самки (Бобринский и др., 1965; Соколов, 1973; Курсков, 1978). Кроме этого можно вспомнить гекконов (виды родов Gymnodactylus, Alsophylax, Crossobamon), откладывающих два крупных яйца в известковой скорлупе и при этом сохраняющих нормальное строение таза (Карр, 1975; Кудрявцев и др., 1991). Наиболее относительно крупное яйцо, составляющее по некоторым данным 20–30 % веса самки, откладывают киви (Apteryx sp.) (Карташев, 1974), несколько меньше этот показатель у некоторых куликов. У перевозчика (Actitis hypoleucos) 4 яйца — 117 % веса самки, у песочников (Calidris minuta, C. subminuta, C. alpina) — 121 %, у ряда других куликов —90–100 %. Крупные яйца у мелких чаек и крачек — 3 яйца до 70 % веса самки, однако такие крупные яйца у птиц — не такое уж частое явление (Дементьев, 1940; Карташев, 1974; Дольник, Дольник, 1982). И самое главное — яйцо при откладке проходит между хвостовыми позвонками и седалищными и лобковыми костями, таким образом расхождение последних в стороны, как и разворот в стороны подвздошных костей может способствовать эволюционному увеличению яиц, но вот разворот последних назад из нижнего или передне-нижнего положения явно сужает этот просвет, в связи с чем и исчезают симфизы седалищной (кроме эму (Dromaius novhaehollandiae)) и лобковой (кроме африканского страуса) костей (Шмальгаузен, 1938; Дементьев, 1940). Таким образом, у вышеупомянутых страусообразных сохраняется закрытый канал для выведения яиц при общем для птиц повороте костей назад. Сохранение симфиза в данном случае можно объяснить небольшими относительно общего размера птицы яйцами, хотя и самыми крупными в абсолютном значении среди современных птиц. Остальные птицы в период яйцекладки увеличивают расстояние между концами разошедшихся лобковых костей, кроме этого, сами кости приобретают несколько большую гибкость и подвижность. Прощупывая концы лобковых костей через брюшную стенку у живой особи, возможно определение физиологического состояния (готова к яйцекладке или нет) не только у домашней птицы (Дементьев, 1940; Вракин, Сидорова, 1984), но и у многих диких видов (Флинт и др.,1986). В некоторых случаях в негнездовой период так можно отличить самцов от самок.
Таким образом, если эволюционное исчезновение симфизов лобковой и седалищной костей можно связать с крупными яйцами, то разворот их назад как будто даже противоречит этому.
Роль своеобразного таза птиц в дыхании, учитывая существенные отличия дыхательного акта птиц и млекопитающих, невозможно оценить без сложных физиологических методик. Особенности дыхания вымерших птицеподобных форм динозавров нам неизвестны, и поэтому мы не беремся подтверждать или опровергать данную версию.
Что касается бипедализма, т. е. передвижения преимущественно или исключительно на задних конечностях, то оно достаточно широко распространено среди современных и вымерших форм млекопитающих (Соколов, 1973) и рептилий (Коуэн, 1982; Кэрролл, 1993). Ни у кого из млекопитающих, как прыгающих на задних ногах с синхронным и асинхронным отталкиванием (кенгуру, тушканчики), так и способных к шаговому передвижению на задних конечностях (некоторые виды тушканчиков) не наблюдается тенденции к изменению формы таза. К непродолжительному бегу на задних конечностях способны некоторые современные ящерицы — василиски (Basiliscus sp.), плащеносная ящерица (Chlamydosaurus kingi), однако и у них таз типично трехлучевого рептильного типа. В то же время у современных крокодилов, несмотря на препубический таз (с направленными вперед лобковыми костями) отсутствует лобковый симфиз (Шимкевич, 1922). Среди вымерших групп рептилий обращают на себя внимание формы, относимые к двум группам динозавров — ящеротазовых (Saurisсhia) и птицетазовых (Ornithisсhia), первые из которых имели типично рептильный таз с направленными вперед и вниз лобковыми костями, образующими в районе симфиза массивное утолщение — “башмак”, хотя у некоторых из них, в первую очередь у двуногих, происходило расширение подвздошной кости и прирастание ее к значительному количеству позвонков. У второй группы подвздошные и лобковые кости отогнуты назад, как у птиц. У некоторых форм подвздошная кость срастается с немногими (4–6) позвонками, у других расширяется и охватывает большее число позвонков. У ряда специализированных форм (особенно более крупных поздних — игуанодонтов, стегозавров) как двуногих, так и четвероногих, развивается направленный вперед предлобковый отросток, который занимает положение, сходное с лобковой костью ящеротазовых. И среди первых, и среди вторых описаны формы, передвигающиеся исключительно в двуногом положении. Некоторые из них, по-видимому, были способны к быстрому и маневренному бегу (Рауп, Стенли, 1974; Коуэн, 1982). Положение отдельных частей позвоночного столба и грудной клетки было сходным с птичьим. Сходством обладала длинная и подвижная шея и легкий ажурный череп, у ряда форм челюсти, по-видимому, одевал клюв (Кэрролл, 1993). Сходство ряда ящеротазовых форм с бегающими птицами было настолько велико, что они получили название Ornithomimidae и Avimimidae. Особенно высокое сходство отмечено в скелете задних конечностей, что вероятнее всего обусловливало и сходство мускулатуры. В то же время среди птицетазовых, несмотря на меньшее общее сходство, встречались, особенно среди “примитивных” ранних, формы, весьма сходные с птицами по строению задних конечностей (Heterodontosaurus, Hypsilophodon), таза (Scelidosaurus, Protoceratops, Homalocephale), часть из которых передвигалась преимущественно или исключительно на задних конечностях, часть использовала исключительно четвероногое передвижение. Несмотря на ряд попыток объяснения подобных различий в строении таза динозавров (Romer, 1956; Galton, 1969; Charig, 1972; Walker, 1977; Santa Luca, 1980, цит. по: Кэрролл, 1993), большинство ученых так или иначе связывают их с локомоцией и реже с объемом брюшной полости (кишечника). Единого мнения так и не сложилось. Однако наличие сходных по габитусу и локомоции форм с разным строением таза само по себе опровергает такую связь (рис. 1). Существенным отличием, правда, всех этих форм, кроме, видимо, Avimimus, является более вертикальное положение конечностей, чем у современных птиц, особенно направленное вниз, а не вперед, бедро, а также значительно меньшая ширина таза, связанная с направлением хотя и широких подвздошных костей косо вниз, а не в стороны, как у птиц, что создавало все-таки заметно меньший объем тазовой полости. Таким образом, связывать характерное положение лобковых и седалищных костей птиц с двуногостью видимо все-таки нет оснований. Как хорошо показал Курзанов (1987), смещению лобковой и седалищной костей сначала вниз, а потом назад не только у птиц, но и у птицеподобных динозавров предшествовало изменение положения бедра и выход его в нормальном (при стоянии) положении вперед от лобковой кости. Это заставило бегающих авимимид изменить расположение и места прикрепления мускулатуры, связанной с бедром, и освободило в значительной степени, а возможно и полностью, лобковую кость от функции прикрепления мускулатуры, связанной с бедром и способствовало дальнейшей возможности изменения ее положения. По-видимому, такая же ситуация сложилась при развитии ранних бегающих предков птиц, хотя палеонтологические остатки, на которых можно было бы показать этот процесс, отсутствуют. Обоснований подобного поворота бедра несколько, но нам кажется, что наиболее существенное — в изменении положения центра тяжести при статической позе, который смещается вперед при уменьшении массы либо укорочении хвоста. С данной проблемой столкнулись как авимимиды, так и птицы (но не орнитомимиды и другие бегающие динозавры с развитым хвостом).
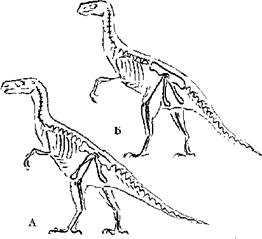
Рис. 1. Строение скелета ящеротазового (А) и птицетазового (Б) динозавров.
Еще меньше вероятность связи птичьего таза с полетом, так как у летающих птерозавров (Pterosauria) таз имел обычное строение (см. также летучие мыши).
Предлагаемое нами объяснение строения таза
Таким образом, рассмотрев предыдущие гипотезы, мы видим, что они не могут быть во всяком случае ведущими в становлении своеобразного для птиц (и ряда динозавров) строения таза. В то же время, вскрывая 1–3-суточных птенцов выводковых птиц, можно обнаружить высокую степень приспособленности данного строения таза с широкими, почти горизонтально поставленными подвздошными костями, широко разнесенными вертлужными впадинами и отодвинутыми назад вверх лобковыми и седалищными костями к вмещению огромного запаса желтка (рис. 2, 3). Этот запас желтка способен поддержать существование птенца иногда до недельного возраста (при оптимальной температуре) без питания (Хейнрот, 1947). Подобный запас желтка, более или менее выраженный, имеют все выводковые птицы. У птенцовых (воробьиные, голуби) он намного меньше, даже у только что вылупившихся птенцов и составляет менее 5 % веса тела (Дементьев, 1940; Хейнрот, 1947; Флинт и др., 1986). Эти различия отражают две экологически разные стратегии эволюции птиц. Типичных птенцовых родители обогревают постоянно и с первых же часов начинают кормить. Запас желтка в такой ситуации практически не нужен, и почти весь желток, находящийся в яйце, еще на ранних стадиях развития идет на построение тела зародыша. Соотношение желтка к белку в яйцах таких птиц 1:2,5 — 1:4 (голуби), 1:3 (дрозды, ласточки), 1:4 (домовый воробей (Passer domesticus)) (Дементьев, 1940). У видов же выводковых соотношение от 1:2 до 1:0,5. При этом часть желтка расходуется в первой трети эмбрионального развития (зародышевый период) на построение тела, а остальная часть (более 2/3) не используется во время эмбрионального роста, когда в предплодном и плодном периоде зародышевого развития эмбрион строит свое тело за счет белка яйца и незадолго перед вылуплением втягивается в брюшную полость, создавая запас на первые дни жизни (Дементьев, 1940; Флинт и др., 1986). Этот запас особенно велик у современных форм, забота родителей о вылупившихся птенцах у которых отсутствует и птенцам приходится самостоятельно обучаться питанию, защитным реакциям и поддерживать температурный баланс. Речь идет о сорных курах (Megapodus eremita, Leipoa ocellata, Telegallus sp.). Соотношение желтка к белку у этих птиц 1:0,5; 1:0,66 соответственно (Хейнрот, 1947). У видов, самостоятельно питающихся с первого дня жизни и находящихся в достаточно жестких температурних условиях, соотношение ниже, например, у гуся — 1:0,9–1:0,97. У видов, первые дни которых проходят в более оптимальных условиях и родители их могут помогать кормодобыванию, соотношение еще ниже: цесарка (Numida meleagris) — 1:1,2; индюк (Meleagris galloopavo), домашняя утка (Anas platyrhynchos) —1:1,4; коростель (Crex crex) — 1:1,6. У курицы (Gallus domesticus), как одомашненного вида, соотношение варьирует — 1:1,7–1:2. У современных птиц в силу широкой адаптивной радиации встречаются все сочетания черт “выводковости” и “птенцовости” вплоть до характерного для чаек и крачек рождения зрячих опушенных и способных к некоторой терморегуляции птенцов, которых все же выкармливают родители до подъема на крыло. Тем не менее, общепринятым считается, что выводковый тип развития для птиц исходен. Наиболее критическим в таких случаях является первый период после вылупления, за который птенцы должны освоить способы поиска и потребления корма и защиты от опасности, хотя в этом им могут помогать родители. При птенцовом типе развития этот критический период исчезает и вся энергия, потребляемая птенцами с кормом, идет на рост тела. У птенцовых развитие и рост идут быстрее. У сходных по размерам и весу сизого голубя (Columbia livia) и серой куропатки (Perdix perdix), стартующих с примерно равных весовых значений, окончательный вес и размер достигается соответственно к 30–40 и 100–120 дню (Поярков, 1955). У наиболее мелких выводковых этот период не меньше 5 недель, у крупных — до 6 месяцев, в то время как достижение окончательных линейных размеров даже у самых крупных птенцовых не более 2 месяцев. У большинства птенцовых более длинный период занимает лишь развитие крупных партий полетного пера (Дементьев, 1940; Чельцов-Бебутов, 1982).
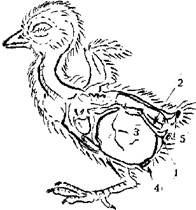
Рис. 2. Размещение запаса желтка у суточного цыпленка:
1 — лобковая кость pubis;
2 — седалищная кость pinbone;
3 — втянутый желток drawn yolk;
4 — пупочное кольцо navel ring;
5 — отверстие клоаки cloacal foramen.
Аналогичный птичьему запас желтка имеется при вылуплении в яйцах черепах и крокодилов, и этот запас тоже расходуется в критический ранний период до перехода на автономное питание. У крокодилов их может хватить иногда на месяц (Даревский, Орлов, 1988), а у черепах, возможно, и на больший срок, но это связано с пойкилотермностью данных видов. У змей и ящериц относительный вес желтка в яйце тоже велик, однако это связано, по-видимому, с тем, что в плодный период развития белок слабо используется для построения тела животного. Запасы желтка при рождении у этих групп незначительны. Высокая постоянная температура тела птиц создает дополнительные сложности и сокращает период, обеспечиваемый питательными веществами втянутого желточного мешка и приводит к необходимости увеличения его объема. У современных крокодилов и черепах при их пропорциях тела как взрослых, так и молоди, не создавалось особых трудностей при помещении значительных объемов остаточного желтка (у черепах внутри панциря, у крокодилов между последними ребрами и отодвинутым назад тазовым поясом). У современных птиц, как и у их ранних форм (насколько мы можем судить по археоптериксу (Archaeopterix lithographica) при коротком туловищном отделе размещение значительных объемов желтка становится сложным, особенно после развития массивной грудины, простирающейся далеко назад. Единственным способом было отодвинуть назад и в стороны элементы тазового пояса. Казалось бы, эта проблема раннего постэмбрионального периода не должна существенно влиять на морфологию взрослых форм, но для птиц как класса в целом одной из характернейших особенностей является детерминированный рост. Пожалуй не найдется ни одной другой группы позвоночных, имеющей такое соотношение размеров новорожденных и взрослых форм (близкое соотношение можно найти только у тоже летающих летучих мышей).
Практически все птицы начинают пользоваться полетом только после достижения линейных размеров взрослой формы. Несколько затушевывают эту важнейшую закономерность несовершенная система промеров птиц, где один из основных — длина крыла с учетом маховых перьев и использование для определения размеров весовых показателей, которые у молодых птиц обычно ниже, чем у взрослых. Хотя есть ряд видов и даже групп, у которых птенцы перед первыми в своей жизни полетами весят больше взрослых за счет больших запасов жира — трубконосые, стрижи, ласточки (Чельцов-Бебутов, 1982). И несколько выпадают из общей закономерности куриные птицы, у которых в связи с особенностями возрастного развития оперения способными к короткому полету оказываются особи, значительно не достигшие окончательных размеров (Хейнрот, 1947). Все остальные виды к началу этапа активного полета оказываются с полностью сросшимися элементами костей, особенно костей крыла, а в большинстве случаев и со значительно пневматизированными участками скелета (Дементьев, 1940; Дольник, 1975; Флинт и др., 1986). По-видимому, продолжение линейного роста и полет — вещи в эволюционном плане несовместимые. Об этом говорят, во-первых, отсутствие линейного роста после начала полета у рукокрылых (Бобринский и др., 1965; Соколов, 1973; Мосияш, 1985) и отсутствие следов линейного роста на костных остатках птерозавров (Кэрролл, 1993).
Второй особенностью птиц как класса является лимитирование у них максимальных, к тому же не очень крупных размеров. Уже давно показана невозможность активного полета у птиц весом более 10–12 кг (Дементьев, 1940; Гладков, 1949; Кокшайский, 1966; Шмидт-Ниельсен, 1978). Мало того, более поздними работами показано, что оптимальный вес для активно летающей птицы лежит в пределах 0,1–1 кг (Кокшайский, 1970; Шмидт-Ниельсен, 1987), а возможно даже в пределах 0,1–0,5 кг (Цвелых, 1983), поскольку более крупным летающим формам приходится формировать специальные морфологические приспособления (увеличение площади крыла, разрезные устройства конца крыла) для покрытия дефицита мощности полетной мускулатуры (Гладков, 1949; Кокшайский, 1966; Якоби, 1966). Птицы весом более килограмма либо редко и несистематически пользуются полетом (дрофы, журавли, куриные), либо переходят к различным типам планирующего полета (трубконосые, аисты, хищники-падальщики), либо полет занимает в их бюджете времени крайне незначительную долю, поскольку энергозатраты на него у таких видов непропорционально велики (Дольник, 1982; Чельцов-Бебутов, 1982). Данное утверждение не касается периода миграций, в течение которого птицы расходуют запасенный в значительных количествах жир и таким образом имеют отрицательный энергобаланс (Дольник, 1975). В целом большинство крупных видов современных птиц относится к высокоспециализированным формам. С другой стороны, наиболее примитивные ранние птицы не могли быть слишком мелкими (типа современных воробьиных) т. е. менее 100 г, т. к. таким формам при полете сложнее преодолевать возникающие завихрения воздуха и ветер (Гладков, 1949; Якоби, 1966; Цвелых, 1983), но, что более важно, у таких мелких современных птиц тоже очень напряженный энергобаланс в связи с противоречием между постоянно поддерживаемой высокой температурой тела и малыми размерами, при которых сохранение тепла превращается в сложную проблему (Шмидт-Ниельсен, 1987; Межжерин, 1988). Таким образом, ранние формы птиц, еще плохо летающие, со слабой полетной мускулатурой, не могли быть слишком крупными, но они не могли быть и слишком мелкими в связи с низкой энерговооруженностью (метаболизм ближе к рептильному, чем к птичьему типу) и, возможно, менее совершенными термоизолирующими качествами покровов.
Следующим существенным моментом для птиц являлась необходимость как можно большего сокращения периода зависимости от родителей (во время гнездования птицы “привязаны” к определенному месту), т. к. до достижения окончательных размеров птицы не могли пользоваться полетом. Все это привело к формированию у птиц, пожалуй, максимально известных среди позвоночных относительных скоростей роста (Шмальгаузен, 1938; Дементьев, 1940; Хейнрот, 1947; Межжерин, 1988). Так, у дроздов, жаворонков, славок линейный рост костных элементов заканчивается иногда к 6–9 дню жизни, и даже у средних по размерам голубей и врановых линейный рост (не учитывая оперения) прекращается к 15–20 дню жизни. Вторым способом сократить период гнездования было увеличение “стартовых значений” для постэмбрионального роста, т. е. увеличение объема и энергетических запасов яйца. Из этого противоречия (крупные размеры яйца и вылупляющегося эмбриона и небольшие окончательные размеры взрослой особи, не меняющиеся с возрастом) сформировалась ситуация, при которой не было времени и “запаса” линейных размеров, за счет которых изменялась бы существенно морфологическая структура, характерная для ранних этапов. В некоторых случаях современные птицы при вылуплении имеют линейные показатели ряда частей тела, характерные для взрослых птиц (лапа некоторых чаек (Larus minutus) и крачек (Sterna hirundo, S. albifrons, род Сhlidonias), цевка ряда мелких чистиковых (Alle, Cepphus, Aethia). Кроме того, увеличенное пространство брюшной — тазовой — полости после использования эмбрионального желтка занималась разросшимся желудком и кишечным трактом у подрастающих особей, а у взрослых позволяла без особого напряжения и “ущемления прав” желудочно-кишечного тракта формировать в яичнике во время размножения крупные яйцеклетки, поскольку при размножении у птицы накладываются друг на друга процессы быстрого роста яйцеклеток в яичнике, формирование белковой и скорлуповой оболочки у находящихся в нижнем отделе яйцевода яиц, причем эти процессы возможны только при интенсивном поступлении питательных веществ и энергии, т. е. опять-таки при максимально эффективной работе пищеварительного аппарата (Дементьев, 1940; Дольник, Дольник, 1982; Флинт и др., 1986). Хорошо известна зависимость яйценоскости, объема яиц, качества желтка и скорлупы и даже ее окраски от интенсивного и сбалансированного питания. Так, например, третьи яйца в кладках чайковых, четвертые — у куликов (группы с детерминированным количеством яиц в кладке) и последние яйца в гнездах воробьиных бледнее и тусклее окрашены, вплоть до совсем белых, и часто мельче по размеру. В то же время, большинство современных рептилий в период роста и формирования яиц, а также на период размножения ограничивают себя в питании либо полностью прекращают питаться, используя накопленные заранее питательные вещества. Крупные размеры эмбрионов у птиц привели к более сложному и дифференцированному развитию в период инкубации и к смещению ряда морфологических изменений на этот период. Так, формирование типичного птичьего таза из возникающей вначале трехлучевой, сходной с рептильной, закладки происходит у куриного, например, эмбриона на восьмой день (Шимкевич, 1922). Втягивание же желтка, как весьма вязкой структуры, происходит в течение нескольких последних дней инкубации и обычно идет параллельно уже начавшемуся легочному дыханию. При этом пупочное отверстие, окруженное кольцевой мускулатурой, имеет значительный диаметр. На этом этапе инкубации довольно велик процент гибели эмбрионов, кроме того, у части их так и не происходит полного втягивания, и пупочное отверстие не закрывается (рис. 3). Такие птенцы, даже нормально вылупившись, без дополнительных хирургических операций обречены на гибель. Таким образом, становится понятным своеобразие темпов роста некоторых структур в постэмбриональном развитии птиц.

Рис. 3. Суточные гусята с втянутым (норма) и невтянутым (нарушение) желтком.
У групп, мало пользующихся полетом во время гнездования (пластинчатоклювые, пастушковые, журавли), особенно при необходимости самостоятельного передвижения и питания, резко отстает в развитии грудина и плечевой пояс, давая возможность на ранних этапах развития увеличить объем тазово-брюшной полости. С этим же связано, видимо, отсутствие существенных отличий по строению и пропорциям таза и грудины между птенцовыми и выводковыми, так как, хотя птенцовым и нет необходимости в размещении остаточного желтка, для них весьма важно на ранних этапах размещения крупного, активно функционирующего желудочно-кишечного тракта и лишь в дальнейшем развитие грудного пояса конечностей, так как они покидают гнездо с помощью полета.
Исходя из всего вышеизложенного мы считаем более соответствующей реальности гипотезу происхождения характерного для птиц строения таза, и в первую очередь отведенные назад лобковые и седалищные кости, первоначально для сохранения на раннем этапе жизни значительных запасов желтка при крупных рождающихся птенцах, жестко лимитированных некрупных размерах взрослой особи, при минимальном времени интенсивного роста.
Происхождение полета
Гипотезы-обоснования
Происхождению полета у птиц и связанных с ним приспособлений посвящено очень много работ, хотя до последнего времени все они так или иначе основываются большей частью на анализе макро- и даже микроскопических признаков археоптерикса (Ostrom, 1978; Feduccia, 1985; Hecht et al., 1985, цит. по: Кэрролл, 1993). У ряда авторов анализируется строение крыла и кисти (анализ этих работ у Курзанова (1987)). Неоднократно анализировался позвоночник, особенно его шейный, очень подвижный у птиц отдел, и строение черепа, обладающего выраженной подвижностью — стрептостилией (анализ у Татаринова (1980) и Кэрролла (1993)) в то время как редукция хвоста и прогрессивное развитие грудины обсуждаются чаще умозрительно, например, Коуэн (1982) Кэролл (1993). Считается доказанным происхождение ранних форм птиц и археоптерикса как по всей вероятности уклоняющейся формы от ящеротазовых предков, так как по новейшим данным (Татаринов, 1980; Кэрролл, 1993) лобковая и седалищные кости у него не заняли еще типично птичьего положения. Многие современные данные говорят о возможном наличии высокого уровня метаболизма у ряда динозавров, по-видимому высокой постоянной температуры тела и как необходимое следствие этого, особенно важное для мелких форм, развитие термоизолирующих покровов. Тем не менее остаются все еще слабо аргументированными возникновение следующих морфологических адаптаций птиц.
1) Наличие летательной поверхности только на передних конечностях, приспособленных в первую очередь для активного гребного полета, и отсутствие подобной поверхности вдоль боков тела, как у всех летающих форм, прошедших стадию планирующего полета. У птиц такая поверхность могла бы формироваться и перьями, как показано, например, на рисунке гипотетического предка у Хейльмана (цит. по: Карташев, 1974); хотя наличие такой перепонки или опахала трудно согласуется с характерным для большинства современных семейств птиц, связанных с древесной растительностью, способом ухода за оперением головы и клювом, производимым задней конечностью через крыло, причем эти способы очень устойчивы в пределах систематических групп.
2) Сохранение на задних конечностях противопоставленного первого пальца и формирование хватательного или лазательного типа задней конечности, естественного и необходимого при прохождении “древесной” стадии, но трудно объяснимого в случае происхождения от быстро бегающих по открытому пространству форм (у современных бегающих птиц и у быстро бегающих вымерших рептилий сокращается число пальцев).
Как известно, к настоящему времени имеется две основные гипотезы происхождения полета у птиц — планирующего полета и прыжка в воздух после длительного бега. Третья, менее распространенная, высказана Остромом (John H. Ostrom, цит. по: Коуэн, 1982). Она связывает начало полета с хлопающими движениями при поимке крупной добычи. Недостатки и достоинства этих гипотез неоднократно обсуждались и приводили в той или иной степени к “гибридизации” их в попытках объяснить трудносовместимые у птиц признаки приспособления к древесному и бегающему образам жизни, что приводит к постулированию “древесной” и “бегающей” стадий. Однако большинство авторов, как ранее, так и в настоящее время, почему-то обсуждают только вопрос происхождения полета “сверху вниз” или “снизу вверх”, совершенно игнорируя тот факт, что полет в первую очередь обеспечивает передвижение на значительные расстояния (как минимум несколько сот длин тела) в горизонтальном направлении, в первую очередь от наземных хищников. Кроме того, как сейчас хорошо из-вестно, взлетный режим в 2–4 раза более энергоемкий, чем устоявшийся горизонтальный полет.
Принимаемые постулаты
Нами при объяснении принимаются следующие основные положения эволюционной теории (Рауп, Стенли, 1974; Северцов, 1981; Грант, 1991; Кэрролл, 1993). Какой-либо признак, особенно приводящий к выраженным морфологическим структурам, на всем протяжении своего формирования должен отвечать определенной функции, хотя возможно и не соответствующей той, которая впоследствии станет для него основной. “Переходные формы” обычно бывают немногочисленными и занимают часто ограниченные районы обитания, в которых происходит эволюция и существенная перестройка функционирования организмов. Только после формирования нового адаптивного типа они могут широко распространяться (гипотеза смены адаптивных зон).
Существенную роль в эволюции, как сито отбора, играют критические периоды жизни вида, которые могут повторяться нерегулярно, иногда раз в десятилетия, и занимать весьма ограниченое время, но приводить к вымиранию 90–98 % популяции с сохранением форм, преадаптированных к данным критическим ситуациям. Хорошие примеры подобного влияния приводит Верещагин (1981, 1986), описывая вымерзание нутрий в Азербайджане и приспособление к нырянию для питания в норме неныряющих речных уток. Весьма часто, если не всегда, группа видов, а чаще вид или даже отдельная популяция, осваивает новую адаптивную зону, не имея особых морфологических приспособлений, только за счет изменения поведения, иногда используя при этом имеющиеся морфологические структуры “не по назначению” (преадаптация) и только в дальнейшем начинает развивать морфологические структуры, более полно приспосабливающие ее к новому способу жизни: защиты от врагов, добывания корма, переживания неблагоприятных условий и т. д. (Дьюсбери, 1981). Кроме этого, в развитии многих морфологических признаков, иногда касающихся разных систем органов, играет большую роль развитие обратных связей, когда развитие какого-либо признака требует наличия соответствующих адаптаций в другой системе органов, но само по себе способствует появлению таких адаптаций.
Предлагаемая нами гипотеза
Первое, что нам необходимо убедительно объяснить — это происхождение перьевого покрова птиц (и, возможно, некоторых птицеподобных форм динозавров). Перьевой покров в развитой форме — великолепный термоизолятор и контурное оперение птиц в сочетании с пухом или хотя бы с пуховой частью контурного пера — сложная структура, требующая достаточно длинного периода совершенствования, на протяжении которого, особенно на ранних стадиях, уже будучи подвижным, но чешуеподобным, не могла участвовать в терморегуляции. По нашему мнению пероподобные чешуйки рептилий, возникших задолго до появления птиц, у некоторых (текодонтов?) использовались для изменения формы (контура) тела, так как сейчас использует ряд современных рептилий раздувание тела (хамелеоны), углов рта (ушастая круглоголовка), специальных воротников и выростов (плащеносная ящерица), горлового мешка (вараны), игуаны (особенно рода Anolis). Подобная практика могла использоваться во внутривидовых демонстрациях либо при попытке напугать врага (Карр, 1975). У современных птиц это широко распространенная форма поведения. Достаточно вспомнить птенцов сов (Asio flammeus, Bubo bubo) в позе угрозы. Подобные чешуйчатые структуры (типа удлиненных чешуек на горле — воротника ящериц рода Lacerta), не неся еще термоизолирующей функции, могли сильно увеличиваться в размерах, дифференцироваться на различных участках тела и привести к “протоперьевому” покрову, который в дальнейшем начал использоваться по своей основной сейчас функции термоизолятора. Этот процесс мог происходить как в линии, приведшей к птицам, так и в других. В это же время, по-видимому, формируется особенность линьки птиц, резко отличающий ее от линьки рептилий. Это касается отдельно выпадения и роста перьев и слущивания тонкого слоя эпидермиса на слабо ороговевающий, хотя и сухой, как у рептилий, коже. Кроме того, быстрый рост и достижение взрослого размера привели к сокращению количества линек; если у рептилий до достижения половозрелости и размеров, близких к взрослым, проходит не менее 8–10, а чаще более 20 линек, то у птиц их число сократилось, по-видимому, до 4–5 (следы этого — два последовательных пуховых покрова, мезоптиль, или промежуточное оперение у некоторых форм и юношеский наряд, практически всегда отличимый от окончательного).
Двуногие формы оперенных мелких рептилий, покрытых уже хорошо функционирующим контурным оперением, но еще без дифференцированных маховых и рулевых, могли обитать в разных биотопах. Одними из наиболее кормных экотонов во все периоды эволюции были прибрежные участки и мелководья, в том числе отк