Мир прокариотной клетки
МИР ПРОКАРИОТНОЙ КЛЕТКИСТРОЕНИЕ ПРОКАРИОТНОЙ КЛЕТКИ
Деления клеточных организмов на высшем уровне предусматривают выделение всех прокариот в отдельное царство. В 70-х гг. обнаружены микроорганизмы, структурно относящиеся к прокариотному типу, но значительно отличающиеся химическим строением важных клеточных макромолекул и способностью осуществлять уникальные биохимические процессы. Эти необычные прокариотные организмы были названы архебактериями. Типичные прокариоты, или бактерии, получили соответственно название эубактерий (истинных бактерий). Число известных архебактерий по сравнению с эубактериями чрезвычайно мало.
Материал настоящего раздела посвящен общей характеристике прокариотных организмов (в основном эубактерий), отличающихся морфологическим и особенно физиологическим разнообразием. В основе морфологического разнообразия лежат различия в размерах и форме отдельных клеток, способах их деления, природе и наборе цитоплазматических включений, строении клеточной стенки и структур, локализованных снаружи от нее, наличии и типе дифференцированных форм, образующихся в процессе жизненного цикла. Всем этим вопросам посвящены главы 4 и 5. В главах 6—9 представлена общая картина физиологического разнообразия прокариот, складывающегося из различий в механизмах получения энергии и источниках питания, разного отношения к молекулярному кислороду и другим факторам внешней среды, прежде всего свету, температуре, кислотности среды. В главе 10 обсуждаются генетические механизмы, приведшие в процессе эволюции к структурно-физиологическому разнообразию прокариот. Глава 11, посвященная проблемам систематики и описанию основных групп прокариот, иллюстрирует на конкретных примерах материал, представленный в предыдущих главах. Завершает раздел глава 12, в которой излагается наиболее общепринятая гипотеза происхождения жизни на Земле, приведшая к возникновению первичной клетки, и имеющийся в настоящее время экспериментальный материал, подтверждающий эту гипотезу.
ФОРМА ПРОКАРИОТ
|
Рис. 3. Разнообразие форм прокариот: 1 — кокк; 2 — диплококк; 3 — сарцина; 4 — стрептококк; 5 — колония сферической формы: 6 — палочковидные бактерии (одиночная клетка и цепочка клеток); 7 — спириллы; 8 — вибрион; 9 — бактерии, имеющие форму замкнутого или незамкнутого кольца; 10 — бактерии, образующие выросты (простеки); 11 — бактерия червеобразной формы; 12 — бактериальная клетка в форме шестиугольной звезды; 13 — представитель актиномицетов; 14 — плодовое тело миксобактерии; 15 — нитчатая бактерия рода Caryophanon с латерально расположенными жгутиками: 16 — нитчатая цианобактерия. образующая споры (акинеты) и гетероцисты; 8, 15, 17, 18 — бактерии с разными типами жгутикования; 19 — бактерии, образующая капсулу; 20 — нитчатые бактерии группы Sphaeroillus , заключенные в чехол, инкрустированный гидратом окиси железа; 21 — бактерия, образующая шипы; 22 — Galionella |
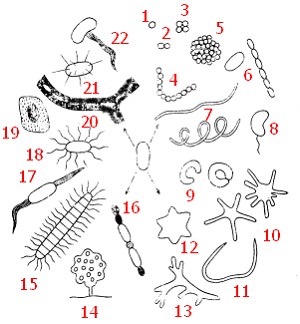
До недавнего времени большинство исследователей традиционно считали, что клетки прокариот достаточно однообразны и в подавляющем большинстве имеют форму сферы, цилиндра или спирали. Они бывают одиночными, в иных случаях образуют нити или колонии. Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (диплококки) или цепочки (стрептококки). В том случае, когда деление происходит относительно равномерно в трех взаимно перпендикулярных направлениях и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы. Если же деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы (рис. 3, 1—5). Прокариоты, имеющие форму цилиндра (палочковидные), сильно различаются по величине отношения длины клетки к ее поперечнику. Прокариоты спиралевидной формы характеризуются разным числом витков: у спирилл — от одного до нескольких витков, вибрионы выглядят наподобие изогнутых палочек, так что их можно рассматривать как неполный виток спирали (рис. 3, 6—8).
За последнее время среди прокариот обнаружены организмы, отличающиеся от описанных выше основных форм. Некоторые бактерии имеют вид кольца, замкнутого или разомкнутого в зависимости от стадии роста (рис.3, 9). У прокариот, в основном размножающихся почкованием, описано образование клеточных выростов (простек), число которых может колебаться от 1 до 8 и более (рис. 3, 10). Из природных субстратов выделены бактерии червеобразной формы и напоминающие шестиугольную звезду (рис. 3, 11, 12). Для некоторых видов характерно слабое или довольно хорошо выраженное ветвление (рис. 3, 13). Описаны прокариоты, обладающие морфологической изменчивостью, в зависимости от условий имеющие вид палочек, кокков или обнаруживающие слабое ветвление.
Форма многоклеточных прокариот также разнообразна: это скопления различной конфигурации, чаще — нити (рис. 3, 14—16). Своеобразие бактериальным клеткам придают жгутики, имеющие различное расположение на клеточной поверхности (рис. 3, 8, 15, 17, 18). а также выделения внеклеточных веществ разной химической природы (рис. 3, 19—22).
СТРУКТУРА, ХИМИЧЕСКИЙ СОСТАВ И ФУНКЦИИ КОМПОНЕНТОВ ПРОКАРИОТНОЙ КЛЕТКИ
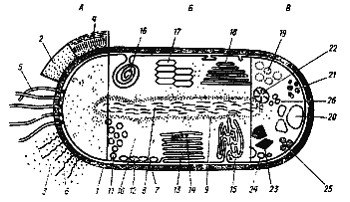
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис. 4). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином "клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур.
|
Рис. 4. Комбинированное изображение прокариотной клетки. А — поверхностные клеточные структуры и внеклеточные образования: 1 — клеточная стенка; 2 — капсула; 3 — слизистые выделения; 4 — чехол; 5 — жгутики; 6 — ворсинки; Б — цитоплазматические клеточные структуры: 7 — ЦПМ; 8 — нуклеоид; 9 — рибосомы; 10 — цитоплазма; 11 — хроматофоры; 12 — хлоросомы; 13 — пластинчатые тилакоиды; 14 — фикобилисомы; 15 — трубчатые тилакоиды; 16 — мезосома; 17 — аэросомы (газовые вакуоли); 18 — ламеллярные структуры; В — запасные вещества: 19 — полисахаридные гранулы; 20 — гранулы поли-b-оксимасляной кислоты; 21 — гранулы полифосфата; 22 — цианофициновые гранулы; 23 — карбоксисомы (полиэдральные тела); 24 — включения серы; 25 — жировые капли; 26 — углеводородные гранулы (по Schlegel, 1972) |
Клеточная стенка
Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50% сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды.
По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна: у так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются 6 . У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
|
Рис. 5. Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликан; 3 — периплазматическое пространство; 4 — наружная мембрана: 5 — цитоплазма, в центре которой расположена ДНК |
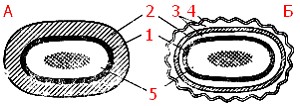
Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу (табл. 3), так и по ультраструктуре (рис. 5).
В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных 1 эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90%), у грамотрицательных — содержание пептидогликана значительно меньше (1—10%). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50% этого гетерополимера.
1 Этот способ был впервые предложен в 1884 г. датским ученым X. Грамом (Ch. Gram), занимавшимся окрашиванием тканей. Позднее он был использован для бактерий.
Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2—3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8—10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
|
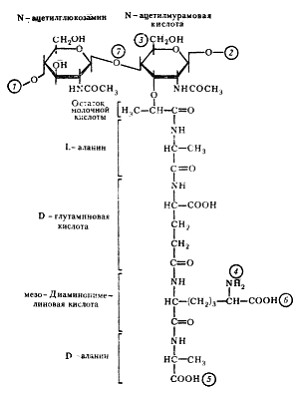
Рис. 6. Структура повторяющейся единицы пептидогликана клеточной стенки эубактерий. Цифры в кружках обозначают: 1, 2— места полимеризации гликанового остова молекулы: 3 — место присоединения с помощью фосфодиэфирной связи молекулы тейхоевой кислоты в клеточной стенке грамположительных эубактерий; 4, 5 — места, по которым происходит связывание между гликановыми цепями с помощью пептидных связей; 6 — место ковалентного связывания (пептидная связь) с липопротеином наружной мембраны у грамотрицательных эубактерий; 7 — место действия лизоцима |
Клеточная стенка грамположительных эубактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм.
Клеточная стенка грамположительных эубактерий. Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер — пептидогликан. Полисахаридный остов молекулы построен из чередующихся остатков N -ацетилглюкозамина и N -ацетилмурамовой кислоты, соединенных между собой посредством) b-1,4-гликозидных связей (рис. 6). К N -ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно 4—5) аминокислот. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы.
Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании дополнительных пептидных связей между гетерополимерными цепочками. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободнаяаминогруппа диаминокислоты другого (рис. 7, А). Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот (рис. 7, Б). Нетрудно себе представить, что этим способом можно "сшить" между собой множество гетерополимерных цепей. Частота "сшивок" различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии.
|
Рис. 7. Пептидные мостики между гетерополимерными цепочками; Г — N -ацетилглюкозамин: М — N -ацетилмурамовая кислота; ала — аланин; глу — глутаминовая кислота; лиз — лизин; ДАП — диаминопимелиновая кислота; гли — глицин. Стрелками обозначено место действия пенициллина |
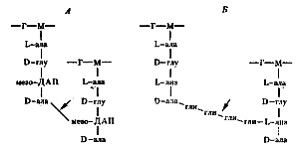
Пептидогликан, окружающий протопласт грамположительных эубактерий, — это по существу одна гигантская молекула, "сшитая" с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей трехмерную пространственную организацию.
Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями (рис. 8). Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N -ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с N -ацетилмурамовой кислотой (см. рис. 6). Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных эубактерий. Остающиеся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона. Как полианионы тейхоевые кислоты определяют поверхностный заряд клетки. Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фага на клеточной поверхности.
|
Рис. 8. Структурная формула глицеринтейхоевой кислоты. Содержит чередующиеся остатки D-аланина и N -ацетилглюкозамина (по Rose, 1971) |
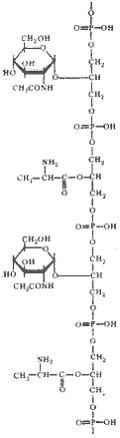
В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки и липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки, в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой.
Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией.
Клеточная стенка бацилл, например Bacillus subtilis , приблизительно соответствует толщине 40 молекул пептидогликана. В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1—6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор.
Клеточная стенка грамотрицательных эубактерий. У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных (см. рис. 5). В ее состав входит гораздо большее число макромолекул разного химического типа (см. табл. 3). Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями (рис. 9).
Таблица 3. Химический состав клеточных стенок грамположительных и грамотрицательных эубактерий (по Rose, 1971; Freer, Salton, 1971)
Компоненты клеточной стенки | Грамположительные эубактерии | Грамотрицательные эубактерии | |
внутренний слой (пептидогликановый) | внешний слой (наружная клеточная мембрана) | ||
Пептидогликан | + | + | – |
Тейхоевые кислоты | + | – | – |
Полисахариды | + | – | + |
Белки | ± | – | + |
Липиды | ± | – | + |
Липополисахариды | – | – | + |
Липопротеины | – | ± | + |
Обозначения: (–) — отсутствуют, (+) — присутствуют, (±) — присутствуют не у всех видов
Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий (см. рис. 6; 7, А). Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (рис. 10, А). Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30—40% ее поверхности и локализованный во внешнем слое (рис. 10, 5).
|
Рис. 9. Однослойная структура пептидогликана. Линиями обозначены гетерополимерные цепочки, образованные чередующимися остатками N -ацетилглюкозамина (Г) и N -ацетилмурамовой кислоты (М), соединенными между собой b-1,4-гликозидными связями. Кружочками обозначены аминокислоты пептидного хвоста |

Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80% всех белков наружной мембраны. Одна из функций этих белков — формирование в мембране гидрофильных пор диаметром примерно 1 нм, через которые осуществляется неспецифическая диффузия молекул с массой до 600—900 Да 7 . Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки наружной мембраны представлены гораздо большим числом видов. Их основная функция — транспортная и рецепторная. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку железосодержащих соединений.
|
Рис. 10. А. Клеточная стенка грамотрицательных эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликановый слой; 3 — периплазматическое пространство; 4 — молекулы белков (заштрихована гидрофобная часть); 5 — фосфолипид; 6 — липополисахарид. Б. Строение молекулы липополисахарида: 1 — липид А; 2 — внутреннее полисахаридное ядро; 3 — наружное полисахаридное ядро; 4 — О-антиген |

Помимо слоев клеточной стенки, типичных для большинства грамотрицательных эубактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной клеточной мембраны. Однако до настоящего времени не ясно, относятся ли они к клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла.
7 Да—дальтон, или единица атомной массы, равен 1,66033x10 -27 кг.
Необычные клеточные стенки прокариот. Некоторые скользящие бактерии (миксобактерии, флексибактерии) способны в процессе перемещения по твердому субстрату периодически менять форму клеток, например путем изгибания, что говорит об эластичности их клеточной стенки, и в первую очередь ее пептидогликанового слоя. Электронно-микроскопическое изучение, однако, обнаружило у них клеточную стенку, типичную для грамотрицательных эубактерий. Наиболее вероятное объяснение гибкости клеточной стенки этих бактерий — чрезвычайно низкая сшитость ее пептидогликанового компонента.
Наконец, обнаружены прокариоты, клеточная стенка которых по структуре и химическому составу резко отличается от описанных выше типов. Они принадлежат к группе архебактерий (см. гл. 17). Клеточные стенки метанобразующих архебактерий содержат пептидогликан особого химического строения. У других представителей этой группы клеточная стенка состоит исключительно из кислого гетерополисахарида, а у некоторых экстремально галофильных, метанобразующих и ацидотермофильных архебактерий — только из белка. Архебактерии с клеточной стенкой белковой природы не окрашиваются по Граму, остальные типы архебактериальной клеточной стенки дают грамположительную реакцию.
Прокариоты без клеточной стенки. При воздействии определенными химическими веществами оказалось возможным получать в лаборатории из разных видов эубактерий формы с частично (сферопласты) или полностью (протопласты) отсутствующей клеточной стенкой. Впервые это обнаружили при действии на бактериальные клетки лизоцимом, ферментом из группы гликозидаз, содержащимся в яичном белке, слезной жидкости и выделяемом некоторыми бактериями. Лизоцим разрывает b-1,4-гликозидные связи в гетерополисахаридной цепи (см. рис. 6), что в конечном итоге может привести к полному удалению пептидогликана из клеточной стенки. Полученные под действием лизоцима сферопласты (из грамотрицательных эубактерий) или протопласты (из грамположительных) принимают сферическую форму и очень чувствительны к внешнему осмотическому давлению. Существовать они могут только в условиях, когда осмотическое давление питательной среды сбалансировано с осмотическим давлением внутри клетки. В благоприятных условиях сферопласты и протопласты проявляют определенную метаболическую активность, но утрачивают способность к размножению.
Прокариоты, не содержащие клеточной стенки, обнаружены и в природе. Это группа микоплазм, сапрофитов и внутриклеточных паразитов растений, животных и человека. Формы, сходные с микоплазмами, были получены также опытным путем с помощью пенициллина, лизоцима и других факторов. Это так называемые L-формы. В благоприятных условиях они обладают метаболической активностью и способностью к размножению. Предполагают, что микоплазмы произошли в результате мутации, нарушившей синтез веществ клеточной стенки, от обычных бактериальных форм аналогично тому, как в экспериментальных условиях получают генетически стабильные L-формы.
Уникальность химического состава клеточной стенки прокариот, ее отличие от таковой эукариот сделали возможным создание и применение лекарственных препаратов, специфически действующих только на прокариотную клеточную стенку. На этом основано действие пенициллина и некоторых других антибиотиков, подавляющих разные этапы синтеза пептидогликана. Пенициллин, например, ингибирует образование связей между пептидными хвостами на этапе "сшивания" полимера, происходящего в клеточной стенке в процессе роста прокариотной клетки (см. рис. 7).
Функции клеточной стенки прокариот. Клеточная стенка прокариот выполняет разнообразные функции: механически защищает клетку от воздействий окружающей среды, обеспечивает поддержание ее внешней формы, дает возможность клетке существовать в гипотонических растворах. В первую очередь в этом "заслуга" пептидогликана.
Структурная дифференцировка клеточной стенки у грамотрицательных эубактерий, приведшая к формированию дополнительного слоя в виде наружной мембраны, значительно расширила круг функций клеточной стенки. Прежде всего это связано с проблемами проницаемости и транспорта веществ в клетку. Наружная мембрана имеет специфические и неспецифические каналы (поры) для пассивного транспорта веществ и ионов, необходимых клетке, т. е. осуществляет функции молекулярного "сита". Наружная мембрана также препятствует проникновению в клетку токсических веществ, что находит отражение в большей устойчивости грамотрицательных эубактерий (сравнительно с грамположительными) к действию некоторых ядов, химических веществ, ферментов и антибиотиков.
Появление у грамотрицательных эубактерий дополнительной мембраны в составе клеточной стенки фактически привело к созданию обособленной полости (периплазматического пространства), отграниченной от цитоплазмы и внешней среды специфическими мембранами и несущей важную функциональную нагрузку.
Периплазматическое пространство, куда погружен пептидогликановый слой. заполнено раствором, в состав которого входят специфические белки, олигосахариды и неорганические молекулы. Периплазматические белки представлены двумя типами: транспортными белками и гидролитическими ферментами. Транспортные белки — это переносчики, связывающиеся с соответствующими субстратами внешней среды и транспортирующие их от наружной мембраны к цитоплазматической.
Было обнаружено также, что многие бактерии способны в больших количествах вырабатывать ферменты (гликозидазы, протеазы, липазы и др.), гидролизующие все типы полимерных молекул. Последними могут быть как молекулы, синтезируемые самой клеткой, так и чужеродные, попавшие в клетку извне. Отрицательные последствия гидролиза собственных молекул (самопереваривание) очевидны. В то же время прокариоты нуждаются в гидролитических ферментах, так как это расширяет круг используемых ими веществ, включая в него полимеры разного типа. Становится понятна необходимость изолирования этих ферментов от цитоплазматического содержимого. Грамположительные эубактерий выделяют гидролитические ферменты во внешнюю среду, у грамотрицательных — они локализованы в периплазматическом пространстве.
Разнообразные функции выполняют макромолекулы, локализованные частично или полностью на внешней стороне клеточной стенки, контактирующей с окружающей средой; это специфические рецепторы для фагов и колицинов; антигены (липополисахарид грамотрицательных эубактерий, тейхоевые кислоты грамположительных); макромолекулы, обеспечивающие межклеточные взаимодействия при конъюгации, а также между патогенными бактериями и тканями высших организмов.
Капсулы, слизистые слои и чехлы
Снаружи клеточная стенка прокариот часто бывает окружена слизистым веществом. Такие образования в зависимости от структурных особенностей получили название капсул, слизистых слоев или чехлов. Все они являются результатом биосинтеза прокариотами органических полимеров и отложения их вокруг клеток.
Под капсулой понимают слизистое образование, обволакивающее клетку, сохраняющее связь с клеточной стенкой и имеющее аморфное строение (см. рис. 3, 19; 4, 2). Если толщина образования меньше 0,2 мкм и, следовательно, оно может быть обнаружено только с помощью электронного микроскопа, говорят о микрокапсуле. Если больше 0,2 мкм, говорят о макрокапсуле. Последнюю можно видеть в обычный световой микроскоп. Для этого препарат просматривают в капле туши, которая не в состоянии проникнуть в капсулу. На темном фоне выделяются клетки, окруженные светлыми зонами. Если же слизистое вещество имеет аморфный, бесструктурный вид и легко отделяется от поверхности прокариотной клетки, говорят о слизистых слоях, окружающих клетку (см. рис. 4, 3).
В отличие от капсул чехлы имеют тонкую структуру. Нередко в них обнаруживают несколько слоев с разным строением (см. рис. 4, 4). Чехлы ряда бактерий, метаболизм которых связан с окислением восстановленных соединений металлов, часто инкрустированы их окислами. Между этими структурами у прокариот обнаружено много переходных форм, так что иногда нельзя четко отграничивать капсулу от слизистых клеточных выделений или капсулу от чехла.
Наличие капсулы зависит от штамма микроорганизма и условий его культивирования. Бактерии, образующие капсулу, могут легко в результате мутации превращаться в бескапсульные формы, что не приводит к какому-либо нарушению клеточной активности, поэтому капсулы нельзя рассматривать как обязательный структурный компонент прокариотной клетки.
Капсулы, слизистые образования и чехлы могут содержать компоненты, одинаковые с клеточной стенкой, однако их химические составы не идентичны. Как правило, химический состав капсул, образуемых бактериями, родо- или видоспецифичен. Основные химические компоненты большинства капсул прокариот — полисахариды гомо- или гетерополимерной природы. Исключение составляет капсула некоторых видов Bacillus, построенная из полипептида, являющегося полимером D-глутаминовой кислоты. Для ряда бактерий показана способность синтезировать и выделять в окружающую среду волокна целлюлозы.
Чехлы как более сложные структуры имеют обычно и более сложный химический состав. Чехол Sphaerotilus natans , например, содержит 36% сахаров, 11 —гексозамина, 27— белка, 5, 2— липида и 0,5% фосфора.
Хотя капсулы, слизистые вещества и чехлы являются необязательными структурами прокариотной клетки, им приписывают определенные полезные для клетки функции. Вязкость внеклеточной среды, обусловленная наличием слизистых веществ, очевидно, благоприятна для клетки. Они защищают клетку от механических повреждений, высыхания, создают дополнительный осмотический барьер, служат препятствием для проникновения фагов. Иногда слизистые образования могут служить источником запасных питательных веществ. С помощью слизи осуществляется связь между соседними клетками в колонии, а также прикрепление клеток к различным поверхностям. Способность определенных бактерий синтезировать эти своеобразные внеклеточные полимеры находит практическое применение: их используют в качестве заменителя плазмы крови, а также для получения синтетических пленок.
Жгутики и механизмы движения
На клеточной поверхности многих прокариот имеются структуры, определяющие способность клетки к движению в жидкой среде. Это — жгутики. Их число, размеры, расположение, как правило, являются признаками, постоянными для определенного вида, и поэтому учитываются при систематике прокариот. Однако накапливаются данные о том, что количество и расположение жгутиков у одного и того же вида могут в значительной степени определяться условиями культивирования и стадией жизненного цикла, и, следовательно, не стоит переоценивать таксономическое значение этого признака.
Если жгутики находятся у полюсов или в полярной области клетки, говорят об их полярном или субполярном расположении, если — вдоль боковой поверхности, говорят о латеральном расположении. В зависимости от числа жгутиков и их локализации на поверхности клетки различают монополярные монотрихи (один жгутик прикреплен к одному полюсу клетки; см. рис. 3, 8), монополярные политрихи (пучок жгутиков расположен на одном полюсе клетки), биполярные политрихи (на каждом полюсе — по пучку жгутиков; см. рис. 3, 17 ) и перитрихи (многочисленные жгутики расположены по всей поверхности клетки или вдоль ее боковой поверхности; см. рис. 3, 18 ). В последнем случае число жгутиков может достигать 1000 на клетку.
|
Рис. 11. Строение жгутика грамотрицательных эубактерий: 1 — нить; 2 — крюк; 3 — базальное тело; 4 — стержень; 5 — L-кольцо; 6 — P-кольцо; 7 — S-кольцо; 8 — M-кольцо; 9 — ЦПМ; 10 — периплазматическое пространство; 11 — пептидогликановый слой; 12 — наружная мембрана (по De Pamphilis, Adier, 1971) |
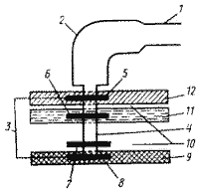
Обычная толщина жгутика — 10—20 нм, длина — от 3 до 15 мкм. У некоторых бактерий длина жгутика может на порядок превышать диаметр клетки. Как правило, полярные жгутики более толстые, чем перитрихиальные. Жгутик представляет собой относительную жесткую спираль, обычно закрученную против часовой стрелки. Вращение жгутика также осуществляется против часовой стрелки с частотой от 40 до 60 об/с, что вызывает вращение клетки, но в противоположном направлении. Поскольку клетка намного массивнее жгутика, она вращается со значительно меньшей скоростью — порядка 12—14 об/мин. Вращательное движение жгутика преобразуется также в поступательное движение клетки, скорость которого в жидкой среде для разных видов бактерий составляет от 16 до 100 мкм/с.
Изучение строения жгутика под электронным микроскопом обнаружило, что он состоит из трех частей (рис. 11). Основную массу жгутика составляет длинная спиральная нить (фибрилла), у поверхности клеточной стенки переходящая в утолщенную изогнутую структуру—крюк. Нить с помощью крюка прикреплена к базальному телу, вмонтированному в ЦПМ и клеточную стенку. У большинства прокариот нить состоит только из одного типа белка — флагеллина. Белковые субъединицы уложены в виде спирали, внутри которой проходит полый канал. Наращивание жгутика происходит с дистального конца, куда субъединицы поступают по внутреннему каналу. У некоторых видов жгутик снаружи дополнительно покрыт чехлом особого химического строения или же являющимся продолжением клеточной стенки и, вероятно, построенным из того же материала.
Крюк (толщина 20—45 нм) состоит из белка, отличающегося от флагеллина, и служит для обеспечения гибкого соединения нити с базальным телом. Базальное тело содержит 9—12 различных белков и представляет собой систему из двух или четырех колец, нанизанных на стержень, являющийся продолжением крюка. Два внутренних кольца (M и S) — обязательные составные части базального тела, в то время как наружные кольца (Р и L) отсутствуют у грамположительных эубактерий и, следовательно, не необходимы для движения. M-кольцо локализовано в ЦПМ, S-кольцо располагается в периплазматическом пространстве грамотрицательных или в пептидогликановом мешке грамположительных эубактерий.
|
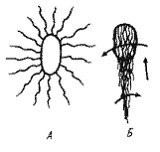
Рис. 12. Клетка Salmonella typhimurium в состоянии покоя (А) и при движении (Б). Стрелками показано направление вращения и движения клетки |
Кольца Р и L, имеющиеся только у грамотрицательных эубактерий, локализованы соответственно в пептидогликановом слое и в наружной мембране. Особенности строения базального тела определяются, таким образом, строением клеточной стенки. Интактность последней необходима для движения жгутиковых бактерий. Обработка клеток лизоцимом, приводящая к удалению пептидогликанового слоя клеточной стенки, вызывает и потерю способности бактерий к движению, хотя жгутики остаются при этом неповрежденными.
Предполагают, что вращение жгутика определяется вращением M-кольца. Другие кольца базального тела неподвижны и служат для крепления стержня, проходящего через клеточную стенку грамотрицательных эубактерий. У грамположительных эубактерий эту функцию в основном выполняет многослойный жесткий пептидогликановый мешок.
Большие успехи достигнуты в расшифровке механизма движения прокариот, имеющих жгутики. Если в клетке много жгутиков. все они при движении собираются в пучок, вращаясь в одном направлении (рис. 12). Вращение жгутиков передается клетке, начинающей вращаться в противоположном направлении, и обеспечивает эффективное движение (плавание) в жидкой среде и более медленное перемещение по поверхности твердых сред.
Для работы двигательного аппарата прокариот необходима энергия. Установлено, что движение жгутиковых прокариот обеспечивается энергией трансмембранного электрохимического потенциала (Dm H + ), причем обе его составляющие — электрическая (Dy) и концентрационная (DpH) — поддерживают движение. Скорость вращения жгутиков прямо зависит от величины мембранного потенциала. Таким образом, прокариотная клетка обладает механизмом, позволяющим превращать электрохимическую форму энергии непосредственно в механическую. Молекулярное устройство, обеспечивающее это превращение, к настоящему времени не выяснено, но можно полагать, что оно должно быть весьма эффективным, так как, по проведенным расчетам, энергия, расходуемая на движение, составляет десятые доли процента от общего количества энергетических потребностей клетки.
|
Рис. 13. Клетка спирохеты в продольном (А) и поперечном (Б) разрезе. На рис. А изображена клетка, содержащая по одной аксиальной фибрилле у каждого конца; на рис. Б — поперечный разрез, прошедший через среднюю часть клетки, где показаны два пересекающихся пучка, состоящих из множества аксиальных фибрилл: 1 — протоплазматический цилиндр; 2 — наружный чехол; 3 — аксиальные фибриллы; 4 — место прикрепления аксиальных фибрилл; 5 — пептидогликановый слой клеточной стенки; 6 — ЦПМ |
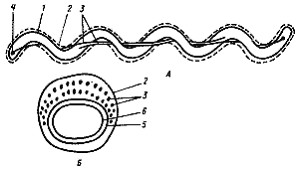
Необычная локализация структур, ответственных за движение, описана у спирохет (рис. 13). Трехслойная структура, окружающая клетку и называемая у спирохет наружным чехлом, аналогична наружной мембране клеточной стенки грамотрицательных эубактерий. Этот чехол окружает так называемый протоплазматический цилиндр, состоящий из пептидогликанового слоя клеточной стенки, ЦПМ и цитоплазматического содержимого. Протоплазматический цилиндр обвивается пучком нитчатых структур — аксиальных фибрилл. Число их колеблется от 2 до 100. Один конец каждой аксиальной фибриллы прикреплен вблизи полюса протоплазматического цилиндра, другой— свободный. Клетка содержит по два набора фибрилл, прикрепленных субполярно у каждого клеточного конца. Так как каждая аксиальная фибрилла тянется почти вдоль всей длины клетки, пучки фибрилл, прикрепленных у разных полюсов, в центральной части перекрываются.
Изучение строения и химического состава аксиальных фибрилл спирохет обнаружило их близкое сходство с бактериальными жгутиками. Отличие заключается в том, что аксиальные фибриллы спирохет — внутриклеточные структуры, но обеспечивают движение как в жидкой среде, так и по твердому субстрату. Движение спирохет осуществляется за счет вращения фибрилл в периплазматическом пространстве между пептидогликановым слоем и наружной мембраной клеточной стенки, вызывающего эластичную волну на поверхности клеточной стенки. Спирохеты совершают движения трех типов: быстро вращаются вокруг длинной оси спирали, способны к изгибанию клеток и осуществляют передвижение по винтовому или волнообразному пути. Для спирохет (так же как для типичных жгутиковых бактерий) показано, что движение обеспечивается энергией в форме Dm H + .
Присущая спирохетам локализация двигательного аппарата интересна тем, что позволяет сделать вывод о возможности его работы в условиях нахождения в "закрытом" клеточными структурами состоянии. Это может служить ключом к пониманию еще одного вида движения, присущего части прокариот, — скольжения. Последнее определяют как способность организма передвигаться по твердому или полужидкому субстрату без помощи наружных локомоторных структур — жгутиков.
Способность к скольжению обнаружена у разных групп прокариот, как одноклеточных, так и многоклеточных, имеющих нитчатое строение: некоторых микоплазм, миксобактерий, цитофаг, нитчатых серобактерий, цианобактерий и др. Скорость этого типа движения невелика: 2–11 мкм/с. Общим для всех скользящих организмов является способность к выделению слизи. Кроме того, у ряда скользящих форм в составе клеточной стенки между пептидогликановым слоем и наружной мембраной обнаружен тонкий слой, состоящий из белковых фибрилл. Например, у нитчатой цианобактерий Oscillatoria к наружной поверхности пептидогликанового слоя примыкают параллельные ряды фибрилл диаметром 5–7 нм: на 1 мкм 2 поверхности приходится до 55 таких фибрилл. У нитчатых цианобактерий фибриллы формируют единую систему, непрерывно в виде спирали обволакивающую весь трихом (нить). Скольжение нитчатых форм сопровождается и одновременным их вращением, так что любая точка на поверхности трихома описывает при движении спираль. Направление вращения является видоспецифическим признаком и коррелирует с направлением хода спирали белковых фибрилл.
Механизм скользящего движения не ясен. Согласно гипотезе реактивного движения оно обусловлено выделением слизи через многочисленные слизевые поры в клеточной стенке, в результате чего клетка отталкивается от субстрата в направлении, противоположном направлению выделения слизи. Однако анализ этой модели привел к заключению, что для обеспечения скольжения по "реактивному" механизму клетке необходимо в течение 1 с выделять такой объем слизи, который во много раз превосходит ее цитоплазматическое содержимое.
По другой гипотезе, получившей распространение в последние годы, скользящее движение связано с особенностями строения клеточной стенки подвижных безжгутиковых форм — наличием белкового слоя, состоящего из упорядоченно расположенных фибрилл, аналогичных нитям жгутиков, с той разницей, что находятся фибриллы "внутри" клеточной стенки. У некоторых скользящих бактерий описаны структуры, весьма напоминающие базальные тела жгутиковых форм. Вращательное движение фибрилл, "запускаемое" этими структурами, приводит к появлению на поверхности клетки так называемой "бегущей волны", т. е. движущихся микроскопических выпуклостей клеточной стенки, в результате чего клетка отталкивается от твердого или вязкого субстрата. На скольжение расходуется около 5% энергии от общего объема клеточных энергетических затрат. Скользящее движение в разных группах бактерий обеспечивается энергией в форме АТФ или Dm H + .
Необходимость для скольжения слизи пока не ясна. Скольжение может происходить в среде подходящей консистенции без какого-либо выделения слизи. Более того, выделение больших количеств слизи, как правило, затрудняет движение клетки и приводит к потере ею подвижности. Согласно гипотезе "бегущей волны" выделение слизи не является абсолютно необходимым для скольжения, но облегчает в определенных условиях отталкивание клетки от субстрата.
Подвижные бактерии активно перемещаются в направлении, определяемом теми или иными внешними факторами. Такие направленные перемещения бактерий называют таксисами. В зависимости от фактора различают хемотаксис (частный случай — аэротаксис), фототаксис, магнитотаксис, термотаксис и вискозитаксис. Наибольшее внимание привлекает изучение хемотаксиса, т. е. движения в определенном направлении относительно источника химического вещества. Для каждого организма все химические вещества в этом плане могут быть разделены на две группы: инертные и вызывающие таксисы (эффекторы). Среди последних выделяют аттрактанты (вещества, привлекающие бактерий) и репелленты (вещества, отпугивающие бактерий). Аттрактантами могут быть сахара, аминокислоты, витамины, нуклеотиды и другие химические молекулы; репеллентами — некоторые аминокислоты, спирты, фенолы, неорганические ионы. Аттрактантом для аэробных и репеллентом для энаэробных прокариот является молекулярный кислород. Аттрактанты часто представлены пищевыми субстратами, хотя не все вещества, необходимые для организма, выступают в качестве аттрактантов. Также не все ядовитые вещества служат репеллентами и не все репелленты вредны.
Фототаксис, т. е. движение к свету или от него, свойствен прежде всего фототрофным бактериям. Способность перемещаться по силовым линиям магнитного поля Земли или магнита — магнитотаксис — обнаружен у разных бактерий, обитающих в пресной и морской воде. В клетках этих бактерий найдены непрозрачные частицы определенной геометрической формы — магнитосомы, заполненные железом в форме магнетита (Fe 3 O 4 ) и выполняющие функцию магнитной стрелки. На долю магнетита может приходиться до 4% сухого вещества бактерий. В северном полушарии такие магниточувствительные бактерии плывут в направлении северного полюса Земли, в южном — в направлении южного. У ряда бактерий обнаружен вискозитаксис — способность реагировать на изменение вязкости раствора и перемещаться в направлении ее увеличения или уменьшения.
За чувствительность бактерий к градиентам определенных факторов ответственны специфические рецепторы. Изучение хемотаксиса у Escherichia coli позволило обнаружить свыше 30 различных хеморецепторов, представляющих собой белки, синтезируемые независимо от присутствия индуктора или только в результате индукции. Рецептор реагирует на эффектор и передает сигнал по определенному пути, конкретный механизм которого неизвестен, на "мотор" жгутика. У бактерий с перитрихиальным жгутикованием выявлены два вида двигательного поведения: прямолинейное движение и кувырканье, т. е. периодические и случайные изменения направления движения. Если бактерия перемещается в сторону оптимальной концентрации аттрактанта, ее прямолинейное движение, ориентированное по отношению к химическому веществу, становится более длительным, а частота кувырканий более низкой, что позволяет ей в конечном итоге перемещаться в нужном направлении.
Ворсинки
К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили) (см. рис. 4, 6). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка — пилина — и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр — 5—10 нм, длина 0,2–2,0 мкм), расположены перитрихиально или полярно. Больше всего сведений имеется о ворсинках Е. coli . У этой бактерии описаны ворсинки общего типа и половые.
Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы.
Наиболее хорошо изучены половые ворсинки, или F-пили, принимающие участие в половом процессе бактерий. F-пили необходимы клетке-донору для обеспечения контакта между ней и реципиентом и в качестве конъюгационного тоннеля, по которому происходит передача ДНК. Ворсинки нельзя считать обязательной клеточной структурой, так как и без них бактерии хорошо растут и размножаются.
Мембраны
Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) — обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8—15% сухого вещества клеток. У большинства прокариотных клеток ЦПМ — единственная мембрана. В клетках фототрофных и ряда хемотрофных прокариот содержатся также мембранные структуры, располагающиеся в цитоплазме и получившие название внутрицитоплазматических мембран. Их происхождение и функции будут рассмотрены ниже.
Химический состав мембран. ЦПМ — белково-липидный комплекс, в котором белки составляют 50–75%, липиды — от 15 до 45%. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95% и больше вещества мембран. Главным липидным компонентом бактериальных мембран являются фосфолипиды — производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 10–30% холестерина, поглощаемого из внешней среды, от общего содержания мембранных липидов. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
|
Рис. 14. Структура основных фосфолипидов мембран эубактерий. R 1 и R 2 — остатки длинноцепочечных жирных кислот, образующих гидрофобный "хвост" молекулы; R 3 может быть остатком глицерина, его производных, этаноламина, инозита и других соединений. Эта часть составляет гидрофильную "голову" молекулы. Простейшим фосфолипидом является фосфатидная кислота, не имеющая R 3 -остатка, связанного с фосфорной кислотой сложноэфирной связью. 1 — общая структура фосфолипида; 2 — фосфатидилглицерин; 3 — дифосфатидилглицерин (кардиолипин); 4 — фосфатидилинозит; 5 — фосфатидилэтаноламин; 6 — фосфатидилсерин |
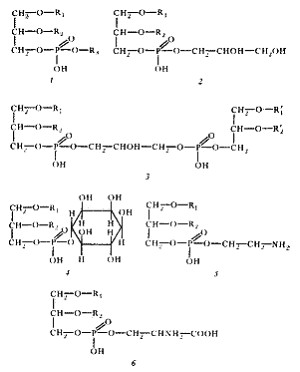
Все липиды эубактерий — производные глицерина — содержат один или несколько остатков жирных кислот, состав которых весьма своеобразен (рис. 14). В основном это насыщенные или мононенасыщенные жирные кислоты с 16—18 углеродными атомами. Полиненасыщенные жирные кислоты у эубактерий отсутствуют. Исключение составляют цианобактерии, у разных видов которых найдены полиненасыщенные жирные кислоты типа C 16:2 , C 18:2 , C 18:3 , C 15:4 . Помимо обычных жирных кислот, т. е. обнаруживаемых и в клетках эукариот, в составе мембранных липидов эубактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Это циклопропановые жирные кислоты, содержащие одно или больше трехчленных колец, присоединенных вдоль углеводородной цепи. Другие, редко встречающиеся и обнаруженные практически только у эубактерий кислоты — это разветвленные жирные кислоты с 15—17 углеродными атомами.
Набор жирных кислот в мембранных липидах также чрезвычайно видоспецифичен. У некоторых грамположительных эубактерий C 15 -жирная кислота с разветвленной цепью может составлять до 90% всех жирных кислот липидов. Главная функция липидов — поддержание механической стабильности мембраны и придание ей гидрофобных свойств.
Особый состав липидов обнаружен в мембранах архебактерий. У них не найдены типичные для эубактерий эфиры глицерина и жирных кислот, но присутствуют эфиры глицерина и высокомолекулярных С 20 -, С 40 -спиртов, а также нейтральные изопреноидные С 20 —С 30 -углеводороды (см. гл. 17).
На долю белков приходится больше половины сухой массы мембран. К мембранам с наиболее высоким содержанием белка относятся бактериальные ЦПМ. При изучении их белкового состава не было обнаружено какого-либо универсального структурного белка. ЦПМ Escherichia coli содержит 27 основных и множество минорных белков, но ни один из основных белков не присутствует в преобладающих количествах. Поскольку ЦПМ прокариот многофункциональна и участвует в осуществлении разнообразных ферментативных процессов, был сделан вывод, что мембранные белки — это, как правило, ферменты. По аминокислотному составу мембранные белки не отличаются от других клеточных белков, за исключением того, что в них содержится мало (иногда следы) цистеина.
В некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. По-видимому, они содержатся не в свободном состоянии, а входят в состав гликолипидов и гликопротеинов.
Структура мембран. Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные "головы" молекул обращены наружу, а гидрофобные "хвосты" погружены в толщу мембраны (рис. 15). Углеводородные цепи, прилегающие к гидрофильным "головам", довольно жестко фиксированы, а более удаленные части "хвостов" обладают достаточной гибкостью. У некоторых архебактерий (ряд метаногенов, термоацидофилы) мембранные липиды, в состав которых входит C 40 -спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойные липидные мембраны обладают большей жесткостью сравнительно с бислойной. При "биологических" температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят в квазикристаллическое состояние. Чем более ненасыщены и разветвлены остатки жирных кислот или чем большее число циклических группировок они содержат, тем ниже температура перехода из жидкостно-кристаллического состояния в квазикристаллическое.
"Жидкая" структура мембран обеспечивает определенную свободу молекул белков, что является необходимым для осуществления процессов транспорта электронов и веществ через мембрану. Это же свойство обусловливает высокую эластичность мембран: они легко сливаются друг с другом, растягиваются и сжимаются.
|
Рис. 15. Модель строения элементарной биологической мембраны: 1 — молекулы липидов: а — гидрофильная "голова"; б — гидрофобный "хвост"; 2 — молекулы белков: в — интегральная; г — периферическая; д — поверхностная. |
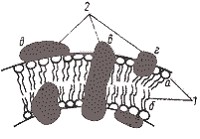
В отличие от липидов у мембранных белков нет единого способа структурной организации. 30–50% белка имеет конфигурацию a-спирали, остальная часть находится преимущественно в виде беспорядочного клубка. Вероятно, часть белков лишена ферментативной активности и участвует только в поддержании мембранной структуры. В то же время доказано, что для осуществления белками некоторых функций необходима их строго упорядоченная взаимная организация в мембране.
В зависимости от расположения в мембране и характера связи с липидным слоем мембранные белки условно можно разделить на три группы: интегральные, периферические и поверхностные (см. рис. 15). Интегральные белки полностью погружены в мембрану, а иногда пронизывают ее насквозь. Связь интегральных белков с мембранными липидами очень прочна и определяется главным образом гидрофобными взаимодействиями. Периферические белки частично погружены в гидрофобную область, а поверхностные находятся вне ее. В первом случае связь с липидами в основном, а во втором — исключительно определяется электростатическими взаимодействиями. Помимо этого некоторые белки и липиды в мембране могут быть связаны ковалентно.
Предложено несколько моделей строения мембраны. Наибольшее признание получила модель, учитывающая большинство данных, известных о мембранах, согласно которой в липидную основу включены асимметрично расположенные белковые молекулы (см. рис. 15). Некоторые из них образуют скопления на поверхностях липидного би- или монослоя, другие частично или полностью погружены в него, третьи пронизывают его насквозь. В модели подчеркнута асимметрия строения мембраны, основанная на различиях в химическом строении и расположении молекул белка.
Функции ЦПМ прокариот. ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ.
Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (Dm H + ), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ является одним из компонентов аппарата генерирования Dm H + . В мембране расположены также ферментные комплексы, обеспечивающие превращения: Dm H + ® АТФ. ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки.
В последнее время выявляется еще одна функциональная грань клеточных мембран — их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией. Клетка — единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода.
ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов 8 . Осуществляется это с использованием разных механизмов мембранного транспорта. Выделяют 4 типа транспортных систем, с участием которых происходит проникновение молекул в бактериальную клетку: пассивную диффузию, облегченную диффузию, активный транспорт и перенос химически модифицированных молекул.
8 У грамположительных форм ЦПМ является и единственным барьером такого рода, у грамотрицательных эубактерий функции дополнительного барьера (молекулярного "сита") выполняет наружная мембрана клеточной стенки, через которую молекулы транспортируются только по механизму пассивной диффузии.
Молекулы воды, некоторых газов (например O 2 , H 2 , N 2 ) и углеводородов, концентрации которых во внешней среде выше, чем в клетке, проходят через ЦПМ внутрь клетки посредством пассивной диффузии. Движущей силой этого процесса служит градиент концентрации вещества по обе стороны мембраны. Основным соединением, поступающим в клетку и покидающим ее таким путем, является вода. Движение воды через мембрану, подчиняющееся законам пассивной диффузии, привело к выводу о существовании в мембране пор. Эти поры пока не удалось увидеть в электронный микроскоп, но некоторые данные о них были получены косвенными методами. Расчетным путем установлено, что поры должны быть очень мелкими и занимать небольшую часть поверхности ЦПМ. Высказывается предположение, что они не являются стабильными структурными образованиями, а возникают в результате временных перестроек молекулярной организации мембраны.
Большинство (если не все) гидрофильных веществ поступает в клетку за счет функционирования систем, в состав которых входят специальные переносчики (транслоказы, или пермеазы), так как скорость физической диффузии этих веществ через гидрофобный слой мембраны очень невелика. Переносчики — вещества белковой природы, локализованные в мембране и характеризующиеся высокой субстратной специфичностью,—связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстрата с одной стороны ЦПМ на другую.
Известен механизм транспорта, получивший название облегченной диффузии, требующий для переноса веществ через мембрану участия транслоказ. Перенос веществ в этом случае происходит по градиенту их концентрации и не требует энергетических затрат. Этот механизм транспорта не получил широкого распространения у прокариот. Основным механизмом избирательного переноса веществ через ЦПМ прокариот является активный транспорт, позволяющий "накачивать" в клетку молекулы и ионы против их концентрационных и электрических градиентов. Этот процесс, так же как и облегченная диффузия, протекает при участии локализованных в ЦПМ переносчиков белковой природы с высокой специфичностью к субстрату, но в отличие от облегченной диффузии для движения против электрохимического градиента требует затрат метаболической энергии. Транспорт такого рода должен быть поэтому сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме.
Во всех описанных выше путях переноса веществ через ЦПМ они поступают в клетку в химически неизмененном виде. У прокариот известны системы транспорта, с помощью которых осуществляется поступление в клетку ряда сахаров, при этом процесс их переноса через мембрану сопровождается химической модификацией молекул. Так происходит, например, поступление в клетки многих прокариот молекул глюкозы, в процессе которого они фосфорилируются.
Внутрицитоплазматические мембраны, прокариот. Выше были отмечены различия между прокариотной и эукариотной клетками в отношении их мембранных систем (см. табл. 1). Отсутствие у прокариот типичных органелл, т. е. структур, полностью отграниченных от цитоплазмы элементарными мембранами, — принципиальная особенность их клеточной организации.
В клетках разных групп прокариот обнаружены мембраны, построенные по принципу элементарной, иные, нежели ЦПМ. Строение, химический состав и функции наружной мембраны грамотрицательных эубактерий описаны ранее. Имеющиеся данные говорят о том, что наружную мембрану можно рассматривать как мембрану другого типа, отличного от ЦПМ. Это касается конкретных аспектов ее строения и функционирования, но не основного принципа организации. Однако наружная мембрана относится к поверхностным структурам клетки эубактерий.
Среди внутрицитоплазматических мембран выделяют несколько видов (табл. 4). Развитая система внутрицитоплазматических мембран характерна для большинства фотосинтезирующих эубактерий. Поскольку было показано, что в этих мембранах локализован фотосинтетический аппарат клетки, они получили общее название фотосинтетических мембран. Все фотосинтетические мембраны (как и все внутриклеточные) — производные ЦПМ, возникшие в результате ее разрастания и глубокого впячивания (инвагинации) в цитоплазму. У некоторых организмов (пурпурные бактерии) фотосинтетические мембраны сохранили тесную связь с ЦПМ, легко обнаруживаемую при электронно-микроскопическом изучении ультратонких срезов клетки. У цианобактерий эта связь менее очевидна. Одни авторы считают, что связь фотосинтетических мембран с ЦПМ у цианобактерий всегда существует, но трудно выявляется, поскольку редко попадает в плоскость среза препарата. По другому мнению, фотосинтетические мембраны цианобактерий — структуры, возникшие первоначально из ЦПМ, но впоследствии отделившиеся от нее и являющиеся в настоящее время автономными клеточными компонентами.
Таблица 4. Мембраны прокариот
Прокариоты | Физиологические группы | Мембраны | ||||
наружная клеточная | цито- плазмати- ческая | внутрицито- плазмати- ческие | ||||
фото- синтети- ческие | мезо- сомальные | прочие | ||||
Грам- положи- тельные | хемотрофы | – | + | – | ± | ±*** |
Грам- отрица- тельные | фототрофы | ±* | + | ±** | ±** | – |
хемотрофы | ±* | + | – | ± | ±**** |
* Отсутствует у архебактерий, клеточная стенка которых построена из белковых субъединиц и не окрашивается по Граму. ** Отсутствуют у зеленых бактерий, цианобактерии Gloeobacter violaceus и экстремально галофильных архебактерий. *** Есть у некоторых метанобразующих архебактерий. **** Сильно развиты у нитрифицирующих, некоторых азотфиксирующих, метанокисляющих бактерий.
Внутрицитоплазматические мембраны фотосинтезирующих эубактерий могут иметь вид трубочек, пузырьков (везикул, хроматофоров) или уплощенных замкнутых дисков (тилакоидов), образованных двумя тесно сближенными мембранными пластинами (ламеллами) (см. рис. 4). Система фотосинтетических мембран очень пластична. Ее морфология и степень развития в клетке определяются многими факторами внешней среды (интенсивностью света, концентрацией кислорода, снабжением клетки питательными веществами), а также возрастными характеристиками культуры.
У прокариот, принадлежащих к разным группам, описаны локальные впячивания ЦПМ, получившие название мезосом (см. рис. 4). Хорошо развитые и сложно организованные мезосомы характерны для грамположительных эубактерий. У грамотрицательных видов они встречаются значительно реже и относительно просто организованы. Мезосомы различаются размерами, формой и локализацией в клетке. Выделяют три основных типа мезосом: ламеллярные (пластинчатые), везикулярные (имеющие форму пузырьков) и тубулярные (трубчатые). Часто можно наблюдать мезосомы смешанного типа: состоящие из ламелл, трубочек и пузырьков. По расположению в клетке различают мезосомы, образующиеся в зоне клеточного деления и формирования поперечной перегородки (септы), мезосомы, к которым прикреплен нуклеоид, и мезосомы, сформированные в результате инвагинации периферических участков ЦПМ.
Существуют разные точки зрения относительно роли мезосом в клетке. Согласно одной из них мезосомы не являются обязательной структурой, а служат только для усиления определенных клеточных функций, увеличивая общую "рабочую" поверхность мембран. Получены данные о том, что с мезосомами связано усиление энергетического метаболизма клеток. Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. Для некоторых грамположительных бактерий обнаружено участие мезосом в секреторных процессах.
Высказывается также предположение, что мезосомы не принимают активного участия в процессах клеточного метаболизма, но выполняют структурную функцию, обеспечивая компартментализацию прокариотной клетки, т. е. пространственное разграничение внутриклеточного содержимого на относительно обособленные отсеки, что создает более благоприятные условия для протекания определенных последовательностей ферментативных реакций. Одновременное существование различных гипотез относительно роли мезосом в прокариотной клетке уже указывает на то, что их функции продолжают оставаться неясными.
Сильно развитая система внутрицитоплазматических мембран, морфологически отличающихся от мезосомальных, описана у представителей трех групп грамотрицательных хемотрофных эубактерий (азотфиксирующих, нитрифицирующих и метанокисляющих), для которых показаны высокая активность дыхания, а также способность метаболизировать растворенные в жидкой среде газообразные соединения.
Цитозоль и рибосомы
Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения.
Рибосомы — место синтеза белка — рибонуклеопротеиновые частицы размером 15—20 нм. Их количество в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80–85% всей бактериальной РНК. Отношение рРНК/белок в рибосомах Е. coli составляет 2:1, у других прокариот оно может быть несколько сдвинуто в сторону преобладания белка. Рибосомы прокариот имеют константу седиментации 705, отчего получили название 70S-частиц. Они построены из двух неодинаковых субчастиц: 305- и 50S-субъединиц 9 . 30S-частица содержит одну молекулу 16S-рРНК и в большинстве случаев по одной молекуле белка более 20 видов. 50S-субъединица состоит из двух молекул рРНК (235 и 55). В ее состав входят более 30 различных белков, также представленных, как правило, одной копией. Большая часть рибосомальных белков выполняет структурную функцию.
9 Обозначения 30S, 50S, 70S — константы седиментации, характеризующие скорость, с которой эти частицы осаждаются в центрифуге при определенных стандартных условиях.
Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами.
Генетический аппарат и репликация хромосомы
|
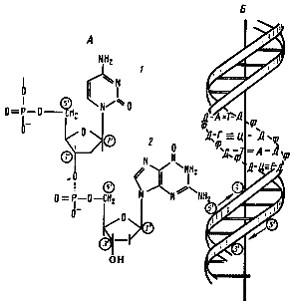
Рис. 16. Строение ДНК: А — фрагмент нити ДНК, образованной чередующимися остатками дезоксирибозы и фосфорной кислоты. К первому углеродному атому дезоксирибозы присоединено азотистое основание: 1 — цитозин; 2 — гуанин; Б — двойная спираль ДНК: Д — дезоксирибоза; Ф — фосфат; А — аденин; Т — тимин; Г — гуанин; Ц — цитозин |
Строение генетического аппарата прокариот долгое время было предметом жарких дискуссий, суть которых сводилась к тому, есть у них такое же ядро, как у эукариот, или нет. Установлено, что генетический материал прокариотных организмов, как и эукариотных, представлен ДНК, но имеются существенные различия в его структурной организации. У прокариот ДНК представляет собой более или менее компактное образование, занимающее определенную область в цитоплазме и не отделенное от нее мембраной, как это имеет место у эукариот. Чтобы подчеркнуть структурные различия в генетическом аппарате прокариотных и эукариотных клеток, предложено у первых его называть нуклеоидом в отличие от ядра у вторых.
При электронно-микроскопическом наблюдении видно, что нуклеоид прокариот, несмотря на отсутствие ядерной мембраны, довольно четко отграничен от цитоплазмы, занимает в ней, как правило, центральную область и заполнен нитями ДНК диаметром около 2 нм. Не исключено, что на выявляемую в электронном микроскопе организацию прокариотной хромосомы большое влияние оказывают условия фиксации препарата. По имеющимся наблюдениям, в живой клетке нуклеоид занимает больше места в цитоплазме.
Вся генетическая информация прокариот содержится в одной молекуле ДНК, имеющей форму ковалентно замкнутого кольца и получившей название бактериальной хромосомы 10 . Длина молекулы в развернутом виде может составлять более 1 мм, т. е. почти в 1000 раз превышать длину бактериальной клетки. Длительное время считали, что в распределении нитей ДНК бактериальной хромосомы не прослеживается никакой закономерности. Однако если исходить из того, что молекула ДНК образует беспорядочный клубок, трудно объяснить процесс репликации и последующее распределение образовавшихся хромосом по дочерним клеткам. Специальные исследования показали, что хромосомы прокариот представляют собой высокоупорядоченную структуру, имеющую константу седиментации 1300—2000S для свободной и 3200—7000S для связанной с мембраной формы. В том и другом случае часть ДНК в этой структуре представлена системой из 20—100 независимо суперспирализованных петель. В обеспечении суперспирализованной организации хромосом участвуют молекулы РНК.
10 В прокариотной клетке ДНК может находиться и вне бактериальной хромосомы — в плазмидах, но последние не являются обязательными клеточными компонентами.
|
Рис. 17. Репликация кольцевой бактериальной хромосомы в двух направлениях. А — родительская молекула ДНК; Б — промежуточные репликативные формы; В — дочерние молекулы ДНК после завершения процесса репликации и расхождения: 1 — точка начала репликации; черными стрелками показано направление репликации |
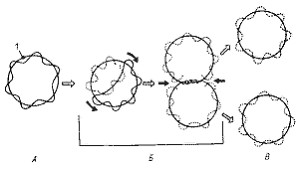
Хромосомы большинства прокариот имеют молекулярную массу в пределах 1—3x10 9 Да. В группе микоплазм генетический материал представлен молекулами, имеющими наименьшее для клеточных организмов количество ДНК (0,4–0,8x10 9 ), а наибольшее содержание ДНК обнаружено у нитчатых цианобактерий (8,5x10 9 ). Хотя каждая прокариотная клетка содержит 1 хромосому, часто в экспоненциально растущей культуре количество ДНК на клетку может достигать массы 3, 4, 8 и более хромосом. Нередко в клетках при действии на них определенных факторов (температуры, pH среды, ионизирующего излучения, солей тяжелых металлов, некоторых антибиотиков и др.) происходит образование множества копий хромосомы. При устранении воздействия этих факторов, а также после перехода в стационарную фазу в клетках, как правило, обнаруживается по одной копии хромосомы.
ДНК прокариот построена так же, как и эукариот (рис. 16). Молекула ДНК несет множество отрицательных зарядов, поскольку каждый фосфатный остаток содержит ионизированную гидроксильную группу. У эукариот отрицательные заряды нейтрализуются образованием комплекса ДНК с основными белками — гистонами. В клетках подавляющего большинства прокариот не обнаружено гистонов, поэтому нейтрализация зарядов осуществляется взаимодействием ДНК с полиаминами (спермином и спермидином), а также с ионами Mg 2+ . В последнее время у некоторых архебактерий и цианобактерий обнаружены гистоны и гистоноподобные белки, связанные с ДНК. Содержание пар оснований А+Т и Г+Ц в молекуле ДНК является постоянным для данного вида организма и служит важным диагностическим признаком. У прокариот молярная доля ГЦ в ДНК колеблется в очень широких пределах: от 23 до 75%.
|
Рис. 18. Механизм распределения бактериальных хромосом: А — бактериальная клетка содержит частично реплицированную хромосому, прикрепленную к мембране в точке (или точках) репликации; Б — репликация хромосомы завершена. В бактериальной клетке две дочерние хромосомы, каждая из которых прикреплена к ЦПМ. Показан синтез клеточной стенки и ЦПМ; В — продолжающийся синтез мембраны и клеточной стенки приводит к разделению дочерних хромосом. Показано начало деления клетки путем образования поперечной перегородки: 1 — ДНК; 2 — прикрепление хромосомы к ЦПМ: 3 — ЦПМ; 4 — клеточная стенка: 5 — синтезированный участок ЦПМ; 6 — новый материал клеточной стенки |
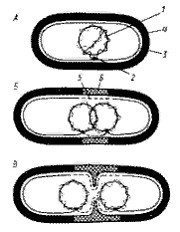
Деление молекулы ДНК (репликация) происходит по полуконсервативному механизму и в норме всегда предшествует делению клетки. С помощью электронного микроскопа установлено, что репликация ДНК начинается в точке прикрепления кольцевой хромосомы к ЦПМ, где локализован ферментативный аппарат, ответственный за репликацию. Часто можно обнаружить, что контакт ДНК с ЦПМ осуществляется посредством мезосом. Репликация, начавшаяся в точке прикрепления, идет затем в двух противоположных направлениях, образуя характерные для кольцевой хромосомы промежуточные структуры (рис. 17). Возникающие дочерние хромосомы остаются прикрепленными к мембране. Репликация молекул ДНК происходит параллельно с синтезом мембраны в области контакта ДНК с ЦПМ. Это приводит к разделению (сегрегации) дочерних молекул ДНК и оформлению обособленных хромосом (рис. 18).
11 Из изложенного выше следует, что термины "нуклеоид" и "хромосома" не всегда совпадают. В зависимости от условий нуклеоид прокариотной клетки может состоять из одной или некоторого числа копий хромосомы.
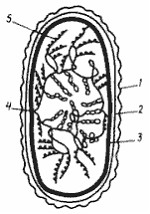
Модель строения бактериальной хромосомы должна объяснять также прохождение в клетке процессов транскрипции и трансляции. Согласно существующим представлениям суперспирализованные петли соответствуют неактивным в данное время участкам ДНК и находятся в центре нуклеоида. По его периферии располагаются деспирализованные участки, на которых происходит синтез информационной РНК (иРНК), при этом, поскольку у бактерий процессы транскрипции и трансляции идут одновременно, одна и та же молекула иРНК может быть одновременно связана с ДНК и рибосомами (рис. 19).
|
Рис. 19. Модель организации нуклеоида Е. coli — наружная мембрана клеточной стенки; 2 — пептидогликановый слой; 3 — ЦПМ; 4 — точка прикрепления бактериальной хромосомы к ЦПМ; 5 — рибосомы, "сидящие" на иРНК. остальные объяснения см. в тексте (по Громову, 1985) |
Рост и способы размножения
Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению.
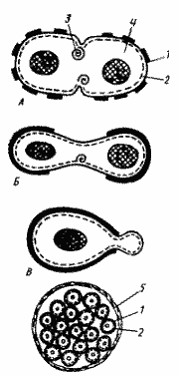
Для подавляющего большинства прокариот характерно равновеликое бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток. При таком способе деления имеет место симметрия в отношении продольной и поперечной оси. У большинства грамположительных эубактерий и нитчатых цианобактерий деление происходит путем синтеза поперечной перегородки, идущего от периферии к центру (рис. 20, А). Так у Bacillus subtilis в середине клетки сначала имеет место кольцевое впячивание ЦПМ, сопровождающееся формированием мезосом разного внешнего вида. Они образуются в месте закладки поперечной перегородки, и предполагается их активное участие в процессах синтеза пептидогликана и других компонентов клеточной стенки. Поперечная перегородка формируется из ЦПМ и пептидогликанового слоя, ее наружные слои синтезируются позднее. Клетки большинства грамотрицательных эубактерий делятся путем перетяжки. У Е. coli на месте деления обнаруживается постепенно увеличивающееся и направленное внутрь искривление ЦПМ и клеточной стенки (рис. 20, 5). Синтез новой клеточной стенки может происходить в нескольких местах или только в зоне формирования поперечной перегородки (рис. 20, А, Б).
Вариантом бинарного деления является почкование, которое можно рассматривать как неравновеликое бинарное деление. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка), увеличивающийся в процессе роста. Постепенно почка достигает размеров материнской клетки, после чего отделяется от последней. Клеточная стенка почки полностью синтезируется заново (рис. 20, В). В процессе почкования симметрия наблюдается в отношении только продольной оси. При равновеликом бинарном делении материнская клетка, делясь, дает начало двум дочерним клеткам и сама, таким образом, исчезает. При почковании материнская клетка дает начало дочерней клетке, и между ними можно в большинстве случаев обнаружить морфологические и физиологические различия: есть старая материнская клетка и новая дочерняя. В этом случае можно наблюдать процесс старения. Так, для некоторых штаммов Rhodomicrobium показано, что материнская клетка способна отпочковывать не более 4 дочерних клеток. Дочерние клетки лучше приспосабливаются к меняющимся условиям. Почкование обнаружено в разных группах прокариот: среди фото- и хемотрофов, осуществляющих авто- и гетеротрофный конструктивный метаболизм. Вероятно, оно в процессе эволюции возникало несколько раз.
|
Рис. 20. Способы деления и синтез клеточной стенки у прокариот: А — деление путем образования поперечной перегородки; Б — деление путем перетяжки; В — почкование; Г — множественное деление: 1 — клеточная стенка (толстой линией обозначена клеточная стенка материнской клетки, тонкой — заново синтезированная); 2 — ЦПМ; 3 — мембранная структура; 4 — цитоплазма, в центре которой расположен нуклеоид; 5 — дополнительный фибриллярный слой клеточной стенки |
Бинарное деление может происходить в одной или нескольких плоскостях. В первом случае, если после деления клетки не расходятся, это приводит к образованию цепочек палочковидных или сферических клеток, во втором — к клеточным скоплениям разной формы (см. рис. 3, 4–6). Расхождение образовавшихся дочерних клеток происходит в результате лизиса среднего слоя клеточной стенки.
Для одной группы одноклеточных цианобактерий описано размножение путем множественного деления. Оно начинается с предварительной репликации хромосомы и увеличения размеров вегетативной клетки, которая затем претерпевает ряд быстрых последовательных бинарных делений, происходящих внутри дополнительного фибриллярного слоя материнской клеточной стенки. Это приводит к образованию мелких клеток, получивших название баеоцитов 12 , число которых у разных видов колеблется от 4 до 1000. Освобождение баеоцитов происходит путем разрыва материнской клеточной стенки (рис. 20, Г ). Таким образом, в основе множественного деления лежит принцип равновеликого бинарного деления. Отличие заключается в том, что в этом случае после бинарного деления не происходит роста образовавшихся дочерних клеток, а они снова подвергаются делению.
Деление прокариотной клетки начинается, как правило, спустя некоторое время после завершения цикла репликации молекулы ДНК. Вероятно, репликация бактериальной хромосомы запускает какие-то процессы, ведущие к клеточному делению. Более детальное изучение у разных видов прокариот взаимосвязи между репликацией ДНК и делением клетки не привело к однозначным результатам. Получены данные о том, что сигналом к клеточному делению служит начало репликации ДНК, ее завершение или репликация определенного локуса бактериальной хромосомы. Таким образом, в норме существует вполне определенная временная связь между репликацией хромосомы и делением бактериальной клетки. Воздействия различными химическими веществами и физическими факторами, приводящие к подавлению репликации ДНК, останавливают и клеточное деление. Однако при некоторых условиях связь между обоими процессами может быть нарушена, и клетки способны делиться в отсутствие синтеза ДНК. Это удалось получить введением определенных мутаций в генетический аппарат бактериальной клетки.
Нарушить последовательность процессов репликации бактериальной хромосомы и клеточного деления также можно, выращивая бактерии при разной температуре. Культивирование Bacillus subtilis на богатой питательной среде при 37° приводит к интенсивному делению бактериальной хромосомы и росту клеток, в результате чего в культуре образуются нитевидные клетки, содержащие множество хромосомных копий с отсутствующими совсем или недосформированными (незамкнутыми) поперечными перегородками. При замедлении скорости роста наблюдается деление нитевидных клеток, приводящее к образованию бактериальных клеток нормальной длины.
Внутрицитоплазматические включения
В цитоплазме прокариот обнаруживаются различные включения. Одни из них следует рассматривать как активно функционирующие структуры, другие — как продукты клеточного метаболизма, не выделяющиеся наружу, но откладывающиеся внутри клетки. Некоторые цитоплазматические включения имеют явно приспособительное значение. И наконец, многие из них являются запасными веществами, отложение которых клеткой происходит в условиях избытка питательных веществ в окружающей среде, а потребление наблюдается, когда организм попадает в условия голодания.
12 Баеоцит — по-гречески маленькая клетка.
К числу внутрицитоплазматических включений, выполняющих определенную функцию в фотосинтезе, относятся хлоросомы зеленых бактерий и фикобилисомы цианобактерий. В этих структурах локализованы пигменты, поглощающие кванты света и передающие их в реакционные центры, т. е. выполняющие роль антенны. Хлоросомы имеют форму продолговатых пузырьков длиной 90—150 и шириной 25—70 нм, окруженных однословной электронно-плотной мембраной толщиной 2—3 нм, построенной только из белка. Они располагаются в непосредственной близости от ЦПМ, плотно к ней примыкая (см. рис. 4). В хлоросомах локализованы бактериохлорофиллы с, d или е . Водорастворимые пигменты белковой природы (фикобили-протеины) цианобактерий содержатся в особых структурах — фикобилисомах, расположенных правильными рядами на внешних поверхностях фотосинтетических мембран и под электронным микроскопом имеющих вид гранул диаметром 28—55 нм (см. рис. 4).
В клетках некоторых прокариот из групп фототрофных и хемолитотрофных эубактерий содержатся структуры, имеющие форму многогранника с 4—6 сторонами и диаметром 90— 500 нм, получившие название карбоксисом, или полиэдральных тел (см. рис. 4). Под электронным микроскопом удалось показать, что они заполнены гранулярным содержимым и окружены однослойной мембраной белковой природы толщиной примерно 3 нм. Карбоксисомы состоят из частиц рибулозодифосфаткарбоксилазы, фермента, катализирующего фиксацию СО 2 на рибулозодифосфате в восстановительном пентозофосфатном цикле. До настоящего времени окончательно не выяснено, в какой форме находится фермент в карбоксисомах: в инертном или функционирующем состоянии. Имеются данные в пользу того, что в активно растущей культуре больше фермента находится в растворимой форме. При переходе в стационарную фазу увеличивается доля рибулозодифосфаткарбоксилазы в составе карбоксисом. Эти данные указывают на возможную роль карбоксисом как структур, обеспечивающих защиту фермента от воздействия внутриклеточными протеазами и, таким образом, его консервирование.
Примером внутрицитоплазматических включений, имеющих приспособительное значение, служат магнитосомы и газовые вакуоли, или аэросомы, обнаруженные у водных прокариот. Газовые вакуоли найдены у представителей, относящихся к 15 таксономическим группам. Это сложно организованные структуры, напоминающие пчелиные соты (см. рис. 4). Состоят из множества регулярно расположенных газовых пузырьков, имеющих форму вытянутого цилиндра с заостренными концами (диаметр 65—115, длина 200—1200 нм). Каждый пузырек окружен однослойной белковой мембраной толщиной 2—3 нм, построенной из одного или двух видов белковых молекул, и заполнен газом, состав которого идентичен таковому окружающей среды. Мембрана газовых пузырьков проницаема для газов, но не проницаема для воды. Число газовых пузырьков, составляющих аэросому, у разных видов различно и зависит от внешних условий. Основная функция газовых вакуолей состоит в обеспечении плавучести водных организмов, которые с их помощью могут регулировать глубину, выбирая более благоприятные условия. При увеличении объема и числа газовых пузырьков плотность цитоплазмы уменьшается, и клетки перемещаются в верхние слои воды. Сжатие газовых пузырьков, наоборот, приводит к погружению клеток. За несколькими исключениями, газовые вакуоли присущи безжгутиковым видам. Их, вероятно, можно рассматривать как альтернативу жгутикам для движения в вертикальной плоскости.
Запасные вещества прокариот представлены полисахаридами, липидами, полипептидами, полифосфатами, отложениями серы (см. рис. 4; табл. 5). Из полисахаридов в клетках откладываются гликоген, крахмал и крахмалоподобное вещество— гранулеза. Последняя — специфический запасной полисахарид анаэробных споровых бактерий группы клостридиев. Названные полисахариды построены из остатков глюкозы. В неблагоприятных условиях они используются в качестве источника углерода и энергии.
Таблица 5. Запасные вещества прокариот
Запасное вещество | Структурные характеристики | Химический состав | Функции | Распространение |
Гранулы гликогена (a-гранулы) | сферической формы, диаметр 20–100 нм | высокомолекулярные полимеры глюкозы | источник углерода и энергии | широко распространенный тип запасных веществ |
Гранулы поли-b-оксимасляной кислоты | диаметр 100–1000 нм; окружены однослойной белковой мембраной 2–3 нм толщиной | 98% полимера поли-b-оксимасляной кислоты, 2% белка | источник углерода и энергии | широко распространены только у прокариот |
Цианофициновые гранулы | размер и форма различны; могут достигать в диаметре 500 нм | полипептид, содержащий аргинин и аспарагиновую кислоту (1:1), мол. масса — 25–100x10 3 Да | источник азота | обнаружены у многих видов цианобактерий |
Гранулы полифосфата | диаметр приблизительно 500 нм, зависит от объекта и условий выращивания | линейные полимеры ортофосфата | источник фосфора и, возможно, энергии | распространенный тип запасных гранул |
Гранулы серы | диаметр 100–800 нм; окружены однослойной белковой мембраной толщиной 2–3 нм | включения жидкой серы | донор электронов или источник энергии | пурпурные серобактерии, бесцветные бактерии, окисляющие H 2 S |
Углеводородные гранулы | диаметр 200–300 нм; окружены белковой оболочкой 2–4 нм толщиной | углеводороды того же типа, что и в среде | источник углерода и энергии | представители родов Arthrobacter , Acinetobacter , Mycobacterium , Nocardia и другие прокариоты, использующие углеводороды |
Липиды накапливаются в виде гранул, резко преломляющих свет и поэтому хорошо различимых в световой микроскоп. Запасным веществом такого рода является полимер b-оксимасляной кислоты, накапливающийся в клетках многих прокариот. У некоторых бактерий, окисляющих углеводороды, поли-b-оксимасляная кислота составляет до 70% сухого вещества клеток. Отложение липидов в клетке происходит в условиях, когда среда богата источником углерода и бедна азотом. Липиды служат для клетки хорошим источником углерода и энергии.
Другой широко распространенный тип запасных веществ многих прокариот — полифосфаты, содержащиеся в гранулах, называемых волютиновыми, или метахроматиновыми, зернами. Используются клетками как источник фосфора. Полифосфаты содержат макроэргические связи и, таким образом, являются депо энергии, хотя считается, что их роль как источника энергии незначительна.
Специфическим запасным веществом цианобактерий являются цианофициновые гранулы. Химический анализ показал, что они состоят из полипептида, содержащего аргинин и аспарагиновую кислоту в эквимолярных количествах. Остов молекулы построен из остатков аспарагиновой кислоты, соединенных пептидными связями, а к ее b-карбоксильным группам присоединены остатки аргинина. Для синтеза цианофицина необходимы затравка, молекулы АТФ, ионы К + и Mg 2+ . Процесс не закодирован в иРНК и не связан с рибосомами. Появление цианофициновых гранул при культивировании цианобактерий в среде с азотом и их исчезновение при истощении среды по азоту указывают на то, что они в клетке служат резервом азота, мобилизуемым при его недостатке в среде.
Для прокариот, метаболизм которых связан с соединениями серы, характерно отложение в клетках молекулярной серы. Сера накапливается, когда в среде содержится сероводород, и окисляется до сульфата, когда весь сероводород среды оказывается исчерпанным. Для аэробных тионовых бактерий, окисляющих H 2 S, сера служит источником энергии, а для анаэробных фотосинтезирующих серобактерий она является донором электронов.
Обращает внимание, что все запасные вещества представлены в виде высокомолекулярных полимерных молекул, в ряде случаев отграниченных от цитоплазмы белковой мембраной, т. е. находятся в осмотически неактивном состоянии. Это важно, так как в противном случае сосредоточение в цитоплазме большого числа молекул осмотически активных веществ оказало бы на клетку отрицательное действие.
ЛИТЕРАТУРА
- Влавианос-Арванитис А., Олескин А.В. Биополитика. Био-окружение. Био-силлабус. Афины: Биополитическая Интернациональная Организация. 1993.
- Горелов А.А. Социальная экология. М. Изд-во Ин-та философии РАН. 1998.
- Гусев М.В. К обсуждению вопроса об антропоцентризме и биоцентризме// Вест. Моск. ун-та. Сер. 16 (Биология). 1991. N 1. С.3--6.
- Данилова Н.Н., Крылова А.Л. Физиология высшей нервной деятельности. М.: Учебная литература. 1997.
- Дерягина М.А. Эволюционная антропология. М.: Изд-во УРАО. 1999.
- Дерягина М.А, Бутовская М.Л. Этология приматов. М.: МГУ. 1992.
- Дольник В.Р. Непослушное дитя биосферы. Беседы о человеке в компании птиц и зверей. М.: Педагогика. 1994.
- Дольник В.Р. Вышли мы все из природы. Беседы о поведении человека в компании птиц, зверей и детей. М.: Linka Press. 1996.
- Дьюсбери Д. Поведение животных. Сравнительные аспекты. М.: Мир. 1981.
- Захаров А.А. Организация сообществ у муравьев. М.: Наука. 1991.
- Зорина З.А., И.И. Полетаева, Ж.И. Резникова. Основы этологии и генетики поведения. М.: Изд-во МГУ. 1999.
- Карпинская Р.С., Лисеев И.К., Огурцов А.П. . Философия природы: коэволюционная стратегия. М.: Интерпракс. 1995. С. 13--78.
- Ламсден Ч., Гуршурст А. Генно-культурная коэволюция: человеческий род в становлении // Человек. 1991. № 3. С.11--22.
- Лоренц К.З. Агрессия (так называемое зло). М.: Прогресс. 1994.
- Майерс Д. Социальная психология. Спб., М., Харьков, Минск: Питер. 2000.
- Мак-Фарленд Д. Поведение животных. Этология и психобиология. М.: Мир. 1988.