Популяции
САНКТ-ПЕТЕРБУРГСКИЙ УНИВЕРСИТЕТ
МВД РОССИИ
КОНТРОЛЬНАЯ РАБОТАПО ЭКОЛОГИИ
на тему: «ПОПУЛЯЦИИ»
Подготовил слушатель:
ФП ФЭК курс I
группа 414
Ежова А.А.
Проверил рецензент:
______________________
______________________
2002г.
ПЛАН
1 Введениеа) понятие популяции
б) свойства популяций
в) факторы динамики численности популяций
г) пространственное размещение популяций
- Заключение
- Использованная литература
ВВЕДЕНИЕ
Все взаимосвязано со всем - гласит первый экологический закон. Значит, и шага нельзя ступить, не задев, а порой и не нарушив чего-либо из окружающей среды. Каждый шаг человека по обычной лужайке - это десятки погубленных микроорганизмов, спугнутых насекомых, изменяющих миграционные пути, а может быть, и снижающих свою естественную продуктивность.
Уже в прошлом веке возникла тревога человека за судьбу планеты, а в текущем столетии дело дошло до кризиса мировой экологической системы из-за возрастания нагрузок на природную среду.
Загрязнение окружающей среды, истощение природных ресурсов и нарушения экологических связей в экосистемах стали глобальными проблемами. И если человечество будет продолжать идти по нынешнему пути развития, то его гибель, как считают ведущие экологи мира, через два-три поколения неизбежна.
Несмотря на принимаемые Российским государством меры по оздоровлению окружающей среды, экологические отношения по-прежнему продолжают развиваться в неблагоприятном для природы и общества направлении:
а) все так же преобладает ведомственный подход, в результате чего каждый экологопользователь эксплуатирует природные богатства, исходя из своих ведомственных интересов;
6) применяется так называемый ресурсовый подход к эколого-пользованию, вследствие чего вне правовой защиты остаются многие экологические связи и природные объекты, не имеющие ресурсовой ценности. А ведь, например, истребление хищных животных, не обладающих ресурсовой ценностью, наносит определенный экологический вред.
Взаимодействие общества и природы перешагнуло грань прежнего равновесия еще в прошлом веке, а в настоящий момент уже невозможно обойтись без надлежащего правового вмешательства в данную сферу. Стало насущным требование разработки специального закона об экологической безопасности России.
В отличии от правовой литературы, где природные объекты исследуются, исходя из их экономической ценности для общества, каждый природный объект должен изучаться в совокупности всех его элементов, которые влияют на жизнедеятельность всей окружающей среды в целом.
В настоящее время, в период надвигающегося экологического кризиса на всей планете, всем ныне живущим необходимо решить задачи перехода от эксплуатации и завоевания Природы к ее сохранению и сотрудничеству с ней. В этих условиях экология человека приобретает особо важное значение, поскольку нормальные условия его существования прямо зависят от того, насколько человек вписывается в природу, способен познать ее законы и творчески использовать их в своей жизни.
Все более интенсивно потребляя природные ресурсы с помощью колоссально возрастающих по своей мощи технических средств, человечество в прогрессирующей форме улучшало условия развития своей цивилизации и своего роста как биологического вида Homo sapiens. Однако, «завоевывая» природу, оно в значительной мере подорвало естественные основы собственной жизнедеятельности, что создало напряженную и во многих случаях кризисную ситуацию во взаимодействии человека и природы, чреватую большими опасностями для будущего цивилизации. Это наглядно обнаруживается, например, при рассмотрении проблем природных ресурсов, энергетики, продовольствия, качества окружающей среды в ее связи с дальнейшим индустриальным развитием в мире и ростом народонаселения. Взаимозависимые изменения привели к возникновению новых связей между глобальной экономикой и глобальной экологией. В прошлом у нас вызывали тревогу последствия экономического роста для окружающей среды. Теперь нас не могут не тревожить последствия «экологического стресса» - ухудшение качества почв, водного режима, состояния атмосферы и лесов - для экономического развития в будущем.
Сейчас становится все более ясным, что источники и причины загрязнения гораздо более разнообразны, сложны и взаимосвязаны, а последствия загрязнения носят более широкий, кумулятивный и хронический характер, чем это считалось ранее. В науке уже даже сформулировано определение антропогенного загрязнения окружающей среды. Это физико-химическое и биологическое изменение качества окружающей среды (атмосферного воздуха, вод, почв) в результате хозяйственной или иной деятельности, превышающее установленные нормативы вредного воздействия на окружающую среду и создающее угрозу здоровью человека, состоянию растительного и животного мира, материальным ценностям.
Экология, как и всякая наука, имеет два аспекта. Один —это стремление к познанию ради самого познания, и в этом плане на первое место ставится поиск закономерностей развития природы, а также их объяснение; другой — применение собранных знаний для решения проблем, связанных с окружающей средой. Стремительное повышение значимости экологии объясняется тем, что ни один из вопросов огромной практической важности в настоящее время нельзя решить без учета связей между живыми и неживыми компонентами природы.
Практический выход экологии можно видеть прежде всего в решении вопросов природопользования; именно она должна создать научную основу эксплуатации природных ресурсов. Мы можем констатировать, что пренебрежение законами, лежащими в основе естественных природных процессов, привело к серьезному конфликту человека и природы. «Совершенно ясно,—пишет американский эколог Р. Риклефс, что никаких очевидных способов исправить нанесенный природе вред не существует, да и обвинение, предъявляемое человеку за его возмутительное отношение к среде, следует строить не на столь очевидных фактах, как сбрасывание в реки сточных вод, опрыскивание посевов пестицидами, ружья и гарпуны охотников, выхлопные газы автомашин, расползающиеся во все стороны пригороды. Человеку следует предъявить обвинение в том, что он не сумел с должным вниманием отнестись к законам, лежащим в основе экономики природы».
ПОНЯТИЕ ПОПУЛЯЦИИ
Объектом изучения демоэкологии, или популяционной экологии, служит популяция. Ее определяют как группу организмов одного вида (внутри которой особи могут обмениваться генетической информацией), занимающую конкретное пространство и функционирующую как часть биотического сообщества. Популяция характеризуется рядом признаков; единственным их носителем является группа, но не особи в этой группе.
Популяция — это совокупность особей одного вида, обитающих на определенной территории, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.
Популяция обладает только ей присущими особенностями: численностью, плотностью, пространственным распределением особей. Различают возрастную, половую, размерную структуру популяции. Соотношение разных по возрасту и полу групп в популяции определяют ее основные функции. Соотношение разных возрастных групп зависит от двух причин: от особенностей жизненного цикла вида и от внешних условий.
Состав. Условно в популяции можно выделить три экологические возрастные группы: пререпродуктивная — группа особей, возраст которых не достиг способности воспроизведения; репродуктивная — группа, воспроизводящая новые особи; пострепродуктивная — особи, утратившие способность участвовать в воспроизведении новых поколений. Длительность этих возрастов по отношению к общей продолжительности жизни сильно варьирует у разных организмов.
Выделяют виды с простой возрастной структурой, когда популяция представлена организмами одного возраста, и виды со сложной возрастной структурой, когда в популяции представлены все возрастные группы или одновременно живут несколько поколений.
Численность и плотность выражают количественные характеристики популяции как целого. Численность популяции выражается числом особей данного вида, обитающих на единице занимаемой ею площади. Динамика численности популяций во времени определяется соотношением показателей рождаемости, смертности, выживаемости, которые в свою очередь определяются условиями жизни.
Плотность популяции — это величина популяции, отнесенная к единице пространства: число особей, или биомасса, популяции на единицу площади или объема. Плотность зависит от трофического уровня, на котором находится популяция. Чем ниже трофический уровень, тем выше плотность.
У многих видов в тех или иных условиях рождаются преимущественно самцы или самки, а иногда особи, неспособные к воспроизведению. У тлей, например, летом сменяют друг друга поколения, состоящие из одних самок. При неблагоприятных условиях появляются самцы. У некоторых брюхоногих моллюсков, многощетинковых червей, рыб, ракообразных происходит изменение пола особи с возрастом.
СВОЙСТВА ПОПУЛЯЦИЙ
От чего же зависят значения коэффициентов рождаемости и смертности? От очень многих факторов, действующих на популяцию извне, а также от собственных ее свойств. Объективный показатель способности организмов увеличивать свою численность —максимальная мгновенная скорость прироста популяции. Этот параметр обратно пропорционален продолжительности жизни организмов. В этом легко убедиться, обратившись к гиперболической зависимости между врожденной скоростью увеличения численности популяции и средним временем генерации, выраженным в днях (рис. 1). Мелкие организмы имеют более высокие значения rтах, чем крупные, что объясняется меньшим временем генерации. Причина подобной корреляции понятна, поскольку организму для достижения крупных размеров требуется много времени. Отсрочка периода размножения также неизбежно ведет к уменьшению rтах.
Тем не менее преимущества, даваемые большими размерами тела, должны превышать недостатки, связанные с уменьшением rтах, так как в противном случае крупные организмы никогда не появились бы в эволюции. Тенденция к возрастанию размеров тела с течением геологического времени, прослеженная по ископаемым остаткам, послужила основанием для введения понятия увеличение филетического размера.
Крупные размеры тела дают совершенно очевидные преимущества: более крупный организм должен привлекать меньше потенциальных хищников и, следовательно, он имеет больше шансов не стать жертвой и должен отличаться лучшей выживаемостью; мелкие организмы находятся в тесной зависимости от физической среды, даже очень слабые изменения которой могут оказаться для них губительными. Более крупные организмы легче переносят воздействие подобных изменений и соответственно лучше от них защищаются. Однако более крупным организмам требуется больше вещества и энергии в расчете на одну особь в единицу времени, чем мелким. Кроме того, для них существует гораздо меньше укрытий и безопасных мест.
В жизни всех организмов в популяции можно выделить три основных периода: предрепродуционный, репродукционный и пострепродукционный. Относительная продолжительность каждого из них у разных видов весьма различна. У многих животных первый период самый длинный. Ярким примером могут служить поденки, у которых предрепродуционный период достигает 3 лет, а репродукционный занимает всего от 2-3 ч до 1 суток. У американской цикады предрепродуционный период составляет 17 лет. Но есть виды, особи которых, едва появившись на свет, начинают интенсивно размножаться (большинство бактерий).
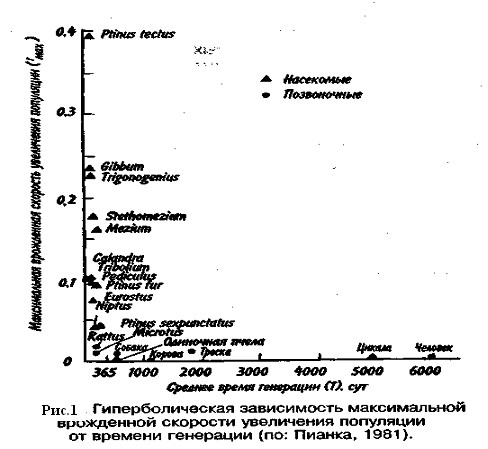
Репродукционные возможности популяции зависят от ее возрастного состава. Продолжительность жизни особей популяции можно оценить, используя кривые выживания. Существует три типа кривых выживания (рис. 2).
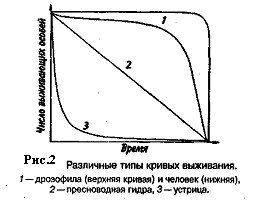
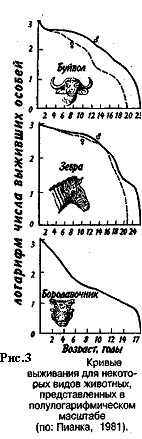
Первый тип (кривые 1) соответствует ситуации, когда большее число особей имеет одинаковую продолжительность жизни и умирает в течение очень короткого отрезка времени. Кривые характеризуются сильно выпуклой формой. Такие кривые выживания свойственны человеку (рис. 2, 1), причем кривая выживания для мужчин по сравнению с аналогичной кривой для женщин менее выпуклая, поэтому страховой полис для мужчин в большинстве стран Запада в 1,5 раза дороже, чем для женщин. Для большинства копытных кривые выживания также выпуклые (рис. 3), хотя и в различной степени для разных видов, а также в зависимости от пола. Второй тип (рис. 2, 2) свойствен видам, коэффициент смертности которых остается постоянным на протяжении всей их жизни. Поэтому кривая выживания трансформируется в прямую линию. Такая форма кривой выживания свойственна пресноводной гидре. Третий тип (рис. 2, 3) — сильно вогнутые кривые, отражающие высокую смертность особей в раннем возрасте. Так характеризуется продолжительность жизни некоторых птиц, рыб, а также многих беспозвоночных.
Знание типа кривой выживания дает возможность построить пирамиду возрастов (рис. 4). Следует различать три типа таких пирамид. Пирамида с широким основанием, что соответствует высокому проценту молодняка, характерна для популяции с большим значением коэффициента рождаемости. Средний тип пирамиды соответствует равномерному распределению особей по возрастам в популяции со сбалансированными коэффициентами рождаемости и смертности — пирамида выровненная. Пирамида с узким основанием (обращенная), отвечающая популяции с численным преобладанием старых особей над молодняком, характерна для сокращающихся популяций. В таких популяциях коэффициент смертности превышает коэффициент рождаемости.

Большое значение для увеличения численности популяции имеют затраты на потомство, выражающиеся в определенной тактике размножения. Не все потомки равноценны: те из них, которые произведены на свет в конце вегетационного сезона, обычно имеют меньшую вероятность дожить до взрослого состояния по сравнению с потомками, появившимися на свет раньше. Сколько усилий должны родители затрачивать на каждого потомка? При постоянной величине репродуктивного усилия средняя приспособленность отдельного потомка связана обратным соотношением с их числом. Один крайний вариант тактики размножения — вложить все в единственного очень крупного и хорошо приспособленного потомка, другой — максимально увеличить общее число произведенных на свет потомков, вложив в каждую особь как можно меньше. Однако наилучшая тактика размножения —это компромисс между производством максимально большого числа потомков и образованием потомства максимально высокой приспособленности.
Указанное соотношение количества и качества потомства иллюстрирует простая графическая модель (рис. 5).
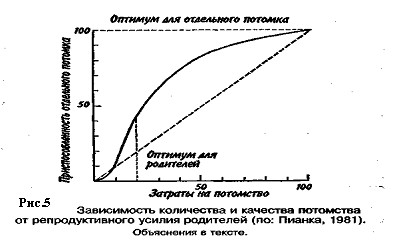
В маловероятном случае, т. е. в случае линейной зависимости приспособленности потомков от затрат на них родителей, приспособленность каждого отдельного потомка уменьшается с увеличением размера помета или кладки. Так как приспособленность родителей или, что то же самое, общая приспособленность всех потомков - постоянная величина, с точки зрения родителей не существует оптимального размера кладки. Однако поскольку первоначальные расходы на потомство вносят больший вклад в приспособленность потомков, чем последующие (имеет место 5-образный характер зависимости приспособленности потомков при увеличении вклада родителей; см. рис. 7.6), то очевидно, что существует некоторый оптимальный размер кладки. В данном гипотетическом случае родители, расходующие только 20% своего репродуктивного усилия на каждого из пяти своих потомков, получат большую отдачу от своего вклада, чем при любом другом размере кладки. Подобная тактика, будучи оптимальной для родителей, не является наилучшей для каждого отдельно взятого потомка, максимальная приспособленность которого достигается в том случае, если он единственный отпрыск, получивший полный вклад усилий от своих родителей. Следовательно, в данном случае налицо «конфликт родителей и детей».
Особенно большое влияние на форму S-образной кривой оказывает конкурентная обстановка. В сильно разреженной среде (конкурентный вакуум) наилучшей репродуктивной стратегией следует считать максимальный вклад вещества и энергии в размножение для продуцирования как можно большего количества потомков в самые короткие сроки. Поскольку конкуренция невелика, потомки могут выжить, даже если они имеют очень малые размеры и низкую приспособленность. Однако в насыщенной среде обитания, где заметно проявляются эффекты массы, а конкуренция остра, оптимальной стратегией будет расходование большого количества энергии на преодоление конкуренции, повышение собственной выживаемости и на продуцирование более конкурентоспособных потомков. При подобной стратегии лучше иметь крупных потомков, а поскольку энергетически они дороже, их может быть произведено на свет меньше.
Итак, свойства популяции можно оценить по таким показателям, как рождаемость, смертность, возрастная структура, соотношение полов, частота генов, генетическое разнообразие, скорость и форма кривой роста и т. д.
Плотность популяции определяется ее внутренними свойствами, а также зависит от факторов, действующих на популяцию извне.
ФАКТОРЫ ДИНАМИКИ ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
Известно три типа зависимости численности популяции от ее плотности (рис. 6). При первом типе (кривая 1) скорость роста популяции уменьшается по мере увеличения плотности. Это широко распространенное явление позволяет понять, почему популяции некоторых животных относительно устойчивы. Во-первых, при увеличении плотности популяции наблюдается снижение рождаемости. Так, в популяции большой синицы при плотности меньше одной пары на 1 га на одно гнездо приходится 14 птенцов; когда же плотность достигает 18 пар на 1 га, выводок составляет менее 8 птенцов. Во-вторых, при увеличении плотности популяции меняется возраст наступления половой зрелости. Например, африканский слон в зависимости от плотности популяции может достигать половой зрелости в возрасте от 12 до 18 лет. Кроме того, этот вид при низкой плотности дает приплод 1 слоненок за 4 года, тогда как при высокой — рождаемость составляет 1 слоненок за 7 лет.
При втором типе зависимости (кривая 2) темп роста популяции максимален при средних, а не при низких значениях плотности. Так, у некоторых видов птиц (например, чаек) число птенцов в выводке увеличивается с повышением плотности популяции, а затем, достигнув наибольшей величины, начинает уменьшаться. Этот тип влияния плотности популяции на скорость размножения особей характерен для видов, у которых отмечен групповой эффект. При третьем типе (кривая 3) темп роста популяции не изменяется до тех пор, пока она не достигнет высокой плотности, затем резко падает.
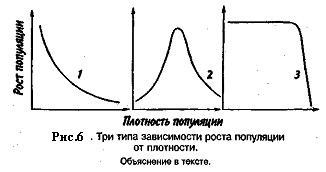
Подобная картина наблюдается, например, у леммингов. При пике численности плотность леммингов становится избыточной, и они начинают мигрировать. Элтон так описал миграции леммингов в Норвегии: животные проходили через деревни в таком количестве, что собаки и кошки, которые вначале нападали на них, просто перестали их замечать. Достигнув моря, обессиленные лемминги тонули.
Регуляция численности равновесных популяций определяется преимущественно биотическими факторами. Среди них главным фактором часто оказывается внутривидовая конкуренция. Примером может служить борьба птиц за мести для гнездования.
Внутривидовая конкуренция может быть причиной физиологического эффекта, известного под названием шоковой болезни. Его отмечают, в частности, у грызунов. Когда плотность популяции становится слишком большой, шоковая болезнь приводит к снижению плодовитости и увеличению смертности, что возвращает плотность популяции к нормальному уровню.
У некоторых видов животных взрослые особи питаются собственным приплодом. Это явление, известное как каннибализм, снижает численность популяции. Каннибализм свойствен, например, окуням: в озерах Западной Сибири 80% пищи крупных особей составляет молодь того же вида. Молодь, в свою очередь, питается планктоном. Таким образом, когда нет других видов рыб, взрослые особи живут за счет планктона.
Межвидовые взаимодействия также играют существенную роль в контроле плотности популяции. Взаимодействия паразит-хозяин и хищник-жертва часто за висят от плотности. Болезни также являются фактором, принимающим участие в регуляции плотности популяции. Когда кролики болеют миксоматозом, вызываемым вирусом, распространение инфекции идет гораздо быстрее в популяциях с повышенной плотностью.
Хищничество как ограничивающий фактор само по себе имеет большое значение. Причем если влияние жертвы на численность популяции хищника не вызывает сомнений, то обратное воздействие, т. е. на популяцию жертвы, бывает не всегда. Во-первых, хищник уничтожает больных животных, тем самым он улучшает средний качественный состав популяции жертвы. Во-вторых, роль хищника ощутима только тогда, когда оба вида обладают приблизительно одинаковым биотическим потенциалом. В противном случае из-за низкого темпа размножения хищник не в состоянии ограничить численность своей жертвы. Например, только одни насекомоядные птицы не могут остановить массовое размножение насекомых. Иными словами, если биотический потенциал хищника намного ниже биотического потенциала жертвы, действие хищника приобретает постоянный характер, не зависящий от плотности его популяции.
Численность насекомых-фитофагов нередко определяется комбинацией видоспецифичных реакций насекомых и растений на воздействие загрязняющих веществ. Загрязнение понижает резистентность растений, в результате чего численность насекомых возрастает. Однако при слишком большом загрязнении численность насекомых падает, несмотря на уменьшение сопротивляемости растений.
Приведенная дифференциация факторов динамики численности популяций позволяет понять их реальное значение в жизни и воспроизводстве популяций. Современная концепция автоматического регулирования численности популяций базируется на сочетании двух принципиально различных явлений: модификаций, или случайных колебаний численности, и регуляций, действующих по принципу кибернетической обратной связи и нивелирующих колебания. В соответствии с этим выделяют модифицирующие (независящие от плотности популяции) и регулирующие (зависящие от плотности популяции) экологические факторы, причем первые из них воздействуют на организмы либо непосредственно, либо через изменения других компонентов биоценоза. По существу, модифицирующие факторы представляют собой различные абиотические факторы. Регулирующие факторы связаны с существованием и активностью живых организмов (биотические факторы), поскольку лишь живые существа способны реагировать на плотность своей популяции и популяций других видов по принципу отрицательной обратной связи (рис. 7).
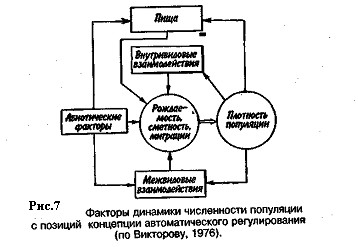
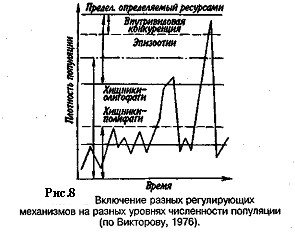
Если воздействия модифицирующих факторов приводят лишь к преобразованиям (модификациям) колебаний численности, не устраняя их, то регулирующие факторы, выравнивая случайные отклонения, стабилизируют (регулируют) численность на определенном уровне. Однако на разных уровнях численности популяции регулирующие факторы принципиально различны (рис. 8). Например, хищники-полифаги, способные при изменении численности жертв ослабить или усилить свою активность—функциональная реакция,— оказывают действие при сравнительно низких значениях численности популяции жертвы.
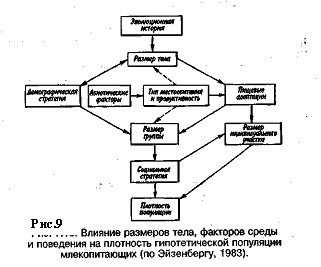
Хищники - олигофаги, характеризующиеся в отличие от полифагов численной реакцией на состояние популяции жертвы, оказывают на нее регулирующее действие в более широком диапазоне, чем полифаги. При достижении популяцией жертвы еще более высокой численности создаются условия для распространения болезней и, наконец, предельный фактор регуляции — внутривидовая конкуренция, ведущая к исчерпыванию доступных ресурсов и развитию стрессовых реакций в популяции жертвы. На рис. 8 представлена многозвенная буферная система регулирования численности популяции под воздействием биотических факторов, степень влияния которых зависит от плотности популяции. В реальной ситуации данный параметр зависит от большого числа факторов, в частности и тех, которые не оказывают регулирующего воздействия на плотность популяции по принципу обратной связи. Взаимодействие между модифицирующими, регулирующими, а также такими специфическими факторами, как размеры тела, группы и индивидуального участка, при их влиянии на плотность популяции млекопитающих показано на рис. 9.
Таким образом, чтобы получить исчерпывающую информацию о том, какие факторы вызывают колебания численности, теоретически требуются данные о физико-химических условиях, обеспеченности ресурсами, жизненном цикле этих организмов и влиянии конкурентов, хищников, паразитов и т. д., причем нужно знать, как все эти факторы влияют на рождаемость, смертность и миграцию. Все популяции непрерывно изменяются: новые организмы рождаются или прибывают как иммигранты, а прежние гибнут или эмигрируют. Несмотря на это, флуктуации размера популяции не безграничны. С одной стороны, она не может расти беспредельно, а с другой — вымирают виды достаточно редко. Следовательно, один из основных признаков популяционной динамики —сочетание изменений с относительной стабильностью. При этом колебания размеров популяций сильно различаются у разных видов.
Особи в популяции взаимодействуют между собой, обеспечивая свою жизнедеятельность и устойчивое воспроизводство популяции.
У животных, ведущих одиночный образ жизни или создающих семьи, регулирующим фактором является территориальность, которая влияет на обладание определенными пищевыми ресурсами и имеет большое значение для размножения. Особь защищает пространство от вторжения и открывает ее для другой особи только при размножении. Самое рациональное использование пространства достигается в том случае, если образуется настоящая территория — участок, из которого другие особи изгоняются. Так как владелец участка психологически господствует на нем, для изгнания чаще всего достаточно лишь демонстрации угроз, преследования, самое большее — притворных атак, которые прекращаются еще на границах участка. У данных животных огромное значение имеют индивидуальные различия между особями — наиболее приспособленные обладают большим индивидуальным пищевым ареалом.
У животных, ведущих групповой образ жизни и образующих стаи, стада, колонии, групповая защита от врагов и совместная за- бота о потомстве повышает выживаемость особей, что влияет на численность популяции и ее выживаемость. Данные животные организованы иерархически. Иерархические отношения соподчинения построены на том, что ранг каждого известен каждому. Как правило, высший ранг принадлежит самому старшему самцу. Иерархия контролирует все взаимодействия внутри популяции: брачные, особей разных возрастов, родителей и потомства.
У животных особую роль играют отношения «мать—дитя». Родители передают потомкам генетическую информацию и информацию о внешней среде.
ПРОСТРАНСТВЕННОЕ РАЗМЕЩЕНИЕ ПОПУЛЯЦИЙ
На уровне популяции абиотические факторы воздействуют на такие параметры, как рождаемость, смертность, средняя продолжительность жизни особи, скорость роста популяции и ее размеры, нередко являясь важнейшими причинами, определяющими характер динамики численности популяции и пространственное распределение особей в ней. Популяция может приспосабливаться к изменениям абиотических факторов, во-первых, меняя характер своего пространственного распределения и, во-вторых, путем адаптивной эволюции.
Избирательное отношение животных и растений к факторам среды порождает избирательность и к местообитаниям, т. е. экологическую специализацию по отношению к участкам ареала вида, которые он пытается занять и заселить. Участок ареала, занятый популяцией вида и характеризующийся определенными экологическими условиями, называется стацией. Выбор стации, как правило, определяется каким-нибудь одним фактором; это может быть кислотность, соленость, влажность и т. д.
Для эвритермных видов характерна зональная смена стаций, т. е. строго направленное изменение стаций при переходе вида из одной природной зоны в другую: при передвижении к северу избираются более сухие, хорошо прогреваемые открытые стации с разреженным растительным покровом, часто располагающиеся на легких песчаных или каменистых почвах; при передвижении к югу тот же вид заселяет более увлажненные и тенистые стации с густым растительным покровом и с глинистыми почвами (рис. 10). На приведенной диаграмме по характеру растительного покрова и микроклимату все стации разбиты на три экологические группы — ксерофитные, мезофитные и гигрофитные. Смещение популяций вида в более влажные стации при продвижении к югу показано косыми стрелками. При этом влаголюбивые популяции лесной и отчасти лесостепной зон лишаются возможности проникнуть в южные районы, так как более влажные, чем гигрофитные, стации физически и экологически немыслимы.
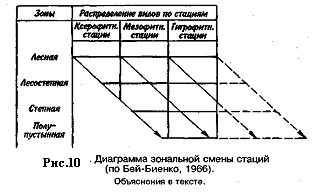
Вертикальная смена стаций аналогична зональной, но проявляется в горных условиях. Наиболее обычной ее формой служит переход популяций на более ксерофитные стации по мере повышения уровня их местообитаний. Например, серый кузнечик в лесном поясе Кавказа держится на мезо- и гигрофитных стациях, а в альпийском поясе — на ксеро- и мезофитных.
Как можно видеть из закономерности смены стаций, важным экологическим фактором, определяющим выбор местообитаний наземных животных и растений, является влажность воздуха.
Специальная приуроченность мокриц связана с содержанием в воздухе водяного пара. Они многочисленны по берегам морей, где воздух насыщен влагой, и там живут открыто. В высокогорных районах с сухим воздухом мокрицы большую часть времени проводят под камнями и корой деревьев.
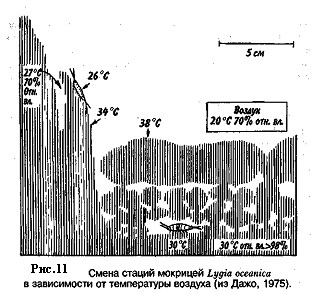
Мокрица Lygia oceanica обитает по берегам морей. Дневное время мокрицы проводят в укрытии. Но когда температура воздуха повышается до 20 °С снаружи и до 30 °С под галькой, они покидают свои укрытия и выползают на скалы, обращенные к солнцу. Причина такого перемещения состоит в том, что данный вид, очень плохо приспособленный к наземному местообитанию, имеет легкопроницаемую кутикулу. Когда влажность воздуха низкая, мокрица теряет много воды путем испарения, что и происходит на скалах, находящихся под солнцем. Интенсивное испарение снижает температуру тела животного, которая при нахождении его на скале равна 26 °С (рис. 11). Если же, мокрица продолжает прятаться под галькой, где относительная влажность близка к 100%, а испарение равно нулю, температура тела достигает 30 °С.
Распределение стаций в водной среде обусловлено другими факторами, в частности кислотностью. Кислые воды торфяников способствуют развитию сфагновых мхов, но в них совершенно нет двустворчатых моллюсков-перловиц. Чрезвычайно редки в них и другие виды двустворчатых моллюсков, что связано с отсутствием в воде извести. Рыбы выносят кислотность воды в пределах рН от 5 до 9. При рН ниже 5 можно наблюдать их массовую гибель, хотя отдельные виды приспосабливаются и к среде, значение рН которой доходит до 3,7. Продуктивность пресных вод, имеющих кислотность менее 5, резко понижена, что влечет за собой значительное сокращение уловов рыбы.
Другой важный фактор, лимитирующий распределение водных животных и растений,—это соленость воды. Многие крупные таксономические группы в ранге типа иглокожие, кишечно-полостные, мшанки, губки, кольчатые черви и др.) все или почти все являются морскими.
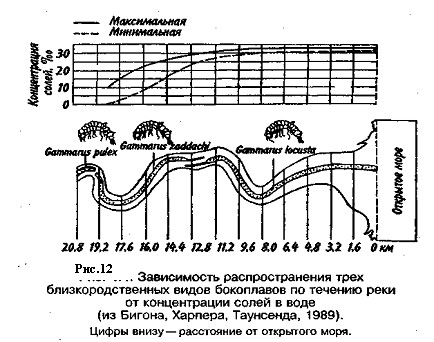
Часто лишь незначительные сдвиги в концентрации солей в воде сказываются на распределении близкородственных видов (рис. 12). Численность обитателей солоноватых вод очень велика, но их видовой состав беден, поскольку здесь могут жить только эвригалинные виды как пресноводного, так и морского происхождения. Например, озеро с соленостью, изменяющейся от 2 до 7%, населяют пресноводные рыбы, такие как карп, линь, щука, судак, хорошо переносящие небольшую соленость, и морские, например кефаль, которые терпимо относятся к недостаточной солености.
Абиотические факторы оказывают существенное влияние на плотность популяций животных и растений. Понижение температуры часто катастрофически сказывается на популяциях животных: в районах, прилегающих к северным границам ареала, вид может стать редким и даже полностью исчезнуть. Кроме того, морозы в ряде случаев влияют и косвенно, поскольку пища, скрытая под толстым слоем льда или снега, становится совершенно недоступной для животных. В местах, подверженных сильным ветрам, рост растений задерживается, а фауна может быть частично или полностью уничтожена.
ЗАКЛЮЧЕНИЕВопрос о том, как эволюционируют экосистемы, очень важен, поскольку его решение — ключ к пониманию существующего разнообразия сообществ живых организмов на нашей планете, смены флоры и фауны в ходе ее геологической истории. В основе эволюции живых организмов лежит естественный отбор, действующий на видовом или более низких уровнях. Но естественный отбор играет также важную роль и на уровне экосистем. Его можно подразделить на взаимный отбор зависящих друг от друга автотрофов и гетеротрофов (коэволюция) и групповой отбор, который ведет к сохранению признаков, благоприятных для экосистемы в целом, даже если они неблагоприятны для конкретных носителей этих признаков.
В самом широком смысле коэволюция означает совместную эволюцию двух (или более) таксонов, которые объединены тесными экологическими связями, но которые не обмениваются генами. Естественный отбор, действующий в популяции хищников, будет постоянно увеличивать эффективность поиска, ловли и поедания добычи. Но в ответ на это в популяции жертвы совершенствуются приспособления, позволяющие особям избежать поимки и уничтожения. Следовательно, в процессе эволюции взаимоотношений «хищник-жертва» жертва действует так, чтобы освободиться от взаимодействия, а хищник—так, чтобы постоянно его поддерживать.
Существуют бесчисленные способы, позволяющие жертвам противостоять давлению хищников. Их можно свести к следующим категориям: защитное поведение (бегство, затаивание, использование убежищ и т. п.), защитная форма и окраска (покровительственная, отпугивающая, предупреждающая, мимикрия), несъедобность или ядовитость (обычно в сочетании с предупреждающей окраской), родительское и социальное поведение (защита потомства, предупреждающие сигналы, совместная защита группы и т. п.). Защитные средства растений включают: жесткие листья, шипы и колючки, ядовитость, репеллентные и ингибирующие питание животных вещества.
Хищники и другие «эксплуататоры» имеют не менее изощренные способы настигнуть жертву. Вспомним, например, общественное охотничье поведение львов и волков, загнутые ядовитые зубы змей, длинные липкие языки лягушек, жаб и ящериц, а также пауков и их паутину, глубоководную рыбу-удильщика или удавов, которые душат свои жертвы.
Животный мир является национальным достоянием России, неотъемлемым элементом природной среды и биологического разнообразия Земли, возобновляющимся природным ресурсом, важным регулирующим и стабилизирующим компонентом биосферы, всемерно охраняемым и рационально используемым для удовлетворения духовных и материальных потребностей народов России.
В ст. 1 Федерального закона о животном мире, принятого Государственной Думой 24 апреля 1995 г. сказано, что животный мир - совокупность живых организмов, всех видов диких животных, постоянно или временно населяющих территорию России, а также относящихся к природным ресурсам континентального шельфа и исключительной экономической зоны Российской Федерации.
Животный мир, являясь составной частью природной среды, выступает как неотъемлемое звено в цепи экологических систем, необходимый компонент в процессе круговорота веществ и энергии природы, активно влияющий на функционирование естественных сообществ, структуру и естественное плодородие почв, формирование растительного покрова, биологические свойства воды и качество окружающей среды в целом, Вместе с тем животный мир имеет большое экономическое значение.
Объектом использования и охраны животного мира выступают лишь дикие животные (млекопитающие, птицы, пресмыкающиеся, земноводные, рыбы, а также моллюски, насекомые и др.), обитающие в состоянии естественной свободы на суше, в воде, атмосфере, в почве, постоянно или временно населяющие территорию России или относящиеся к естественным богатствам континентального шельфа и экономической зоны РФ. Не являются таким объектом сельскохозяйственные и другие домашние животные, а также дикие животные, содержащиеся в неволе или полуневоле для хозяйственных, культурных, научных, эстетических и иных целей. Они являются имуществом, принадлежащим на праве собственности государству, юридическим или физически лицам и используются и охраняются в соответствии с гражданским законодательством.
Особенностью животного мира является то, что данный объект возобновляем, но для этого необходимо соблюдение определенных условий, непосредственного связанных с охраной животных. При истреблении, нарушении условий их существования определенные виды животных могут окончательно исчезнуть, и их возобновление будет невозможно.
Указанный Федеральный закон регулирует отношения в области охраны и использования объектов животного мира (любые виды, популяции, живые организмы, относящиеся к животному миру) и среды их обитания в целях сохранения целостности естественных сообществ, биологического разнообразия и обеспечения устойчивого использования и воспроизводства объектов животного мира, а также для укрепления законности и правопорядка в указанной области.
В Федеральном законе предусмотрены традиционные методы охраны и использования объектов животного мира. Лица, чье существование и доходы полностью или частично основаны на традиционных системах жизнеобеспечения, включая охоту, рыболовство и собирательство, имеют право на применение традиционных методов добывания объектов животного мира и продуктов жизнедеятельности, если такие методы прямо или косвенно не ведут к снижению биологического разнообразия, не сокращают численность и устойчивое воспроизводство объектов животного мира, не нарушают среду их обитания и не представляют опасности для человека. Это право указанные лица могут осуществлять как индивидуально, так и коллективно, создавая объединения на различной основе (семейные, родовые, территориально-хозяйственные общины, союзы охотников, собирателей, рыболовов и иные).
ИСПОЛЬЗОВАННАЯ ЛИТЕРАТУРА
- Краткий курс общей экологии, А.К. Бродский, СПб, «ДЕАН», 1999
- 1000 экзаменационных билетов и готовых ответов.9 кл., Москва, «ДРОФА», 2001
- Экологическое право, Б.В. Ерофеев, Москва, «НОВЫЙ ЮРИСТ», 1998